 2007, Vol. 43
2007, Vol. 43文章信息
- 康华靖, 刘鹏, 陈子林, 廖承川, 李成惠, 陈卫新, 雷祖培.
- Kang Huajing, Liu Peng, Chen Zilin, Liao Chengchuan, Li Chenghui, Chen Weixin, Lei Zupei.
- 不同生境香果树种群的径级结构与分布格局
- Size-Class Structure and Distribution Pattern of Emmenopterys henryi in Different Habitats
- 林业科学, 2007, 43(12): 22-27.
- Scientia Silvae Sinicae, 2007, 43(12): 22-27.
-
文章历史
- 收稿日期:2006-05-08
-
作者相关文章



2. 大盘山国家级自然保护区管理局 磐安 322300;
3. 九龙山国家级自然保护区管理局 遂昌 323300;
4. 乌岩岭国家级自然保护区管理局 泰顺 325500
2. Administration Bureau of Dapanshan Natural Reserve Pan'an 322300;
3. Administration Bureau of Jiulongshan Natural Reserve Suichang 323300;
4. Administration Bureau of Wuyanlin Natural Reserve Taishun 325500
种群是构成群落的基本单位,其结构不仅对群落结构有直接影响,并能客观地体现出群落的发展、演变趋势(Wu et al., 2002),其径级结构能很好地反映种群动态变化(Arista,1995)。种群的分布格局是由种群本身生物学特性、生境条件以及2者之间的相互作用所决定的,是对种群个体在水平空间上配置和分布状态做出的定量描述(闫淑君等,2002;Quinn et al., 1983)。研究种群分布格局的目的不仅是对种群的水平结构进行定量描述,更重要的是用以揭示格局的成因,阐述种群及群落的动态变化等,为保护措施的制定提供科学依据(Greig-Smith,1952)。
香果树(Emmenopterys henryi)属茜草科(Rubiaceae)香果树属(Emmenopterys),为中国特有单种属植物,是研究茜草科系统发育、形态演化及中国植物地理区系的重要材料。由于香果树现存数量有限,濒临灭绝,故被列为国家Ⅱ级重点保护植物(于永福,1999)。目前,国内外对香果树的研究主要集中在种子生理特性(李铁华等,2004;甘聃等,2006)、群落结构特征(徐小玉等,2002)、生物学特性(刘军,2003)、育苗技术(谢玉芳等,2004)、组织培养(韦小丽等,2005)以及遗传多样性(李钧敏等,2006;熊丹等,2006)等方面,但有关不同生境下香果树种群的径级结构和分布格局的研究未见报道。本文通过对不同生境下香果树种群的径级结构和分布格局进行研究,旨在了解香果树种群的更新规律,探讨香果树濒危的机制,进而为合理地保护这一濒危物种提供理论依据。
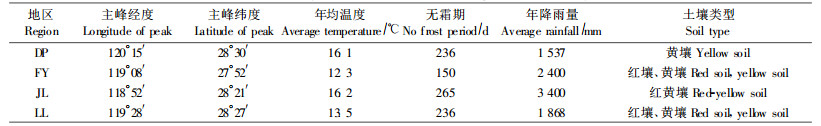
1 研究地概况2005年6月至11月,对浙江省内国家级自然保护区及其林场的香果树分布进行实地考察,选取分布比较集中的大盘山国家级自然保护区(DP)、凤阳山国家级自然保护区(FY)、九龙山国家级自然保护区(JL)和松阳箬寮自然保护区(LL)作为研究地,对香果树种群进行调查统计。根据文献(郝朝运等,2004;杨旭等,2005;王昌腾,2005)和当地气象部门以及相关资料了解的上述5个地区的概况见表 1。
|
|
香果树主要分布于海拔400~1200 m的沟谷水溪旁的山坡上,其伴生乔木种常有杉木(Cunn inghamia lanceolata)、红脉钓樟(Lindera rubronervia)、拟赤扬(Alniphyllum fortunei)、灯台树(Cornus controversa)、柳杉(Cryptomeria fortunei)等,灌木层常有马银花(Rhododendron ovatum)、尖连蕊茶(Camellia cuspidata)、荚迷(Viburnum farreri)、山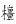
在大盘山国家级自然保护区内选取了4个典型样区,每个样区中设置1个样地,其样地面积为20 m×20 m, 该区内香果树的主要木本伴生种有杉木、红脉钓樟、山NFDA3和尖连蕊茶等;在凤阳山国家级自然保护区(FY)内选取了2个典型样区,每个样区中设置2个样地,其样地面积为15 m×20 m, 该区内香果树的主要木本伴生种有灯台树、红脉钓樟和山NFDA3等;在九龙山国家级自然保护区(JL)也选取了2个典型样区,每个样区设置2个样地,样地面积为20 m×20 m, 该区内香果树的主要木本伴生种有灯台树、拟赤杨等;在松阳箬寮自然保护区(LL)选取了1个典型样区设置了2个样地,样地面积为20 m×20 m, 该区内的香果树主要木本伴生种有柳杉、尖连蕊茶、马银花等。对样地内香果树进行每木调查,详细记录胸径、树高、冠幅、每株香果树的坐标(x,y)等指标。统计时,以香果树基株为单位进行统计,一是种子萌发实生苗长成的植株,二是由根系或树桩萌孽生成的植株。如果一个树桩上有多个无性系分株,则仅对其中径级最大的一株进行记录。同时,记录各样地的海拔、坡向、坡度、坡位、群落郁闭度、岩石裸露度等生态因子(表 2)。
|
|
按照以下标准将径级结构分为5级,标准为:Ⅰ级幼苗<0.33 m;Ⅱ级幼树>0.33 m,胸径(DBH)<2.5 cm;Ⅲ级小树DBH为2.5~7.5 cm;Ⅳ级中树DBH为7.5~22.5 cm;Ⅴ级大树DBH>2 2.5 cm。
2.3 种群分布格局 2.3.1 格局规模分析为了避免单一尺度格局分析的缺陷,采用Greig-Smith法(Gittins,1985)和Kershaw法(Kershaw et al.,1985)分别在1、2、4、8、16、32和64 m2尺度下对香果树种群的分布格局进行分析。
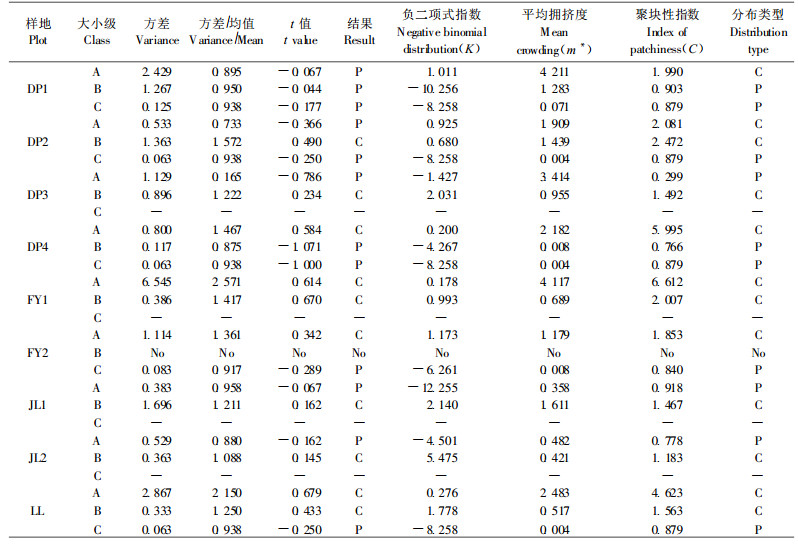
2.3.2 种群分布格局分析把各样地划分成5 m ×5 m的小样方,应用相邻格子法记录样地内的数据资料,用以种群空间分布格局的拟合。离散分布理论拟合采用方差/均值的t检验方法;聚集强度指标采用负二项式分布K、Lloyd平均拥挤度指数m*和聚块指数C(刘建泉,2004)。
2.3.3 不同发育阶段种群分布格局分析将香果树按A(DBH<7.5 cm)、B(DBH为7.5~22.5 cm)、C(DBH>22.5 cm)级的划分标准进行处理,然后按照2.3.2方法进行分析。
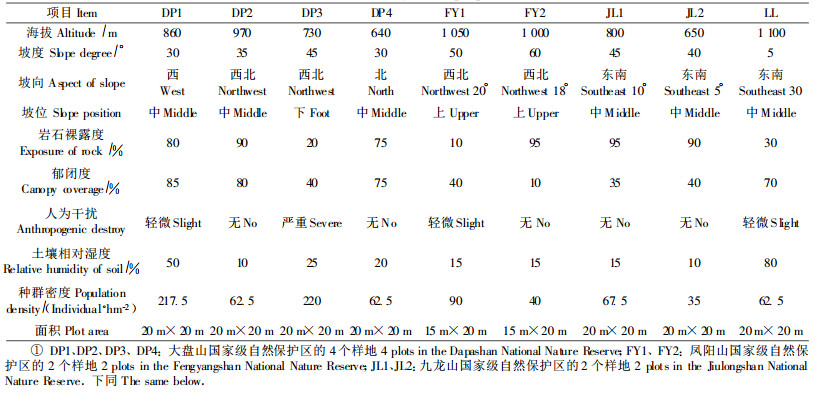
3 结果与分析 3.1 种群径级结构从图 1可以看出,不同生境下香果树种群的年龄结构非常相似,Ⅲ级小树或Ⅳ级中树的数量均较多,而幼苗和幼树较为缺乏,在DP4中甚至没有发现幼苗。由于缺乏幼苗和幼树,香果树种群呈衰退趋势。

|
图 1 不同生境香果树的径级结构 Fig. 1 Size-class structure of E.henryi in different habitats |
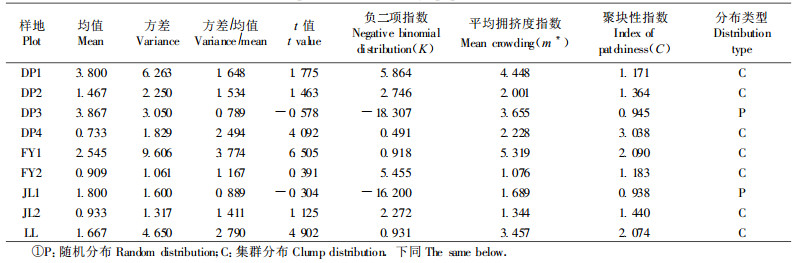
对香果树种群空间分布格局进行分析,从表 3可以看出,Greig-Smih的值一般比较小,特别是FY2、JL2和LL。这与样地内香果树数量偏少有密切的关系。不同生境下样地的Greig-Smith和Kershaw均在8、16或32 m2区组尺度上出现一个较小的峰值,表明香果树的聚块规模为8~32 m2。因此,5 m× 5 m的调查小样方能够比较客观地反映出香果树种群格局规模。
|
|

Kershaw等(1985)的研究结果表明,当个体平均密度极低或极高时,t检验的可靠性有所下降,而K值检验则不受样方内个体密度的影响,故本文主要依据K值进行分布格局的判定(表 4)。从表 4来看,不同生境下的种群分布格局表现出一定的相似性:JL1和DP3为随机分布,其余样地均为集群分布。在DP3中母树被过度砍伐,产生大量的萌生苗,使得该种群呈随机分布;在JL1中群落郁闭度不高,岩石裸露度很高(达到95%左右),也没有树木被破坏,从而使种子和萌枝更新都无法很好地进行,导致种群密度较小,种群也呈随机分布;其余样地内由于裸岩、林木天然死亡、被砍伐和沟溪而形成许多大小不等的林窗,香果树个体为了寻求光照而在林窗内和林窗边缘形成群聚,导致了香果树种群的集群分布。从m*值看,DP1、DP3、FY1和LL样地的平均拥挤度明显高于其他样地,这与样地生境优越,人为干扰及萌枝大量形成有关。
|
|
从表 5可以看出,在DP3、JL1和JL2中A级树木是随机分布,而其他样地的A级树木均为集群分布。这与种群的生境条件以及人为干扰密切相关:DP3生境优越,人为干扰严重,产生大量的萌枝;而JL1和JL2由于岩石裸露度极高,种群保存较为完好,不利于香果树幼苗的生长和萌枝的形成,故A级树木呈随机分布。C级树木(无大树的样地除外)均为随机分布,因此,从A级到C级的时间序列上看,香果树分布格局为从集群到随机,显示出扩散的趋势。
|
|
对不同生境下香果树种群径级结构研究表明,不同生境下的种群均表现出幼龄株数少,老龄株数多的径级结构。这主要由于香果树具有始花期长(刘军,2003)、种子有休眠特性、寿命短(李铁华等,2004)等生物学特性,使得在自然条件下,香果树通过种子更新非常困难;由于香果树种群生境条件比较险恶,多位于水沟旁或岩石裸露度比较高的山坡上,常有水流冲刷地面,所以萌条更新也较为困难。因此,不同生境下的香果树种群均呈衰退趋势。
种群的空间分布格局不但因种而异,而且同一种在不同发育阶段、不同的生境条件和不同尺度上均有明显的差别(江洪,1992)。对不同生境下香果树种群的分布格局研究发现,不同生境下香果树种群的格局规模为8~32 m2。其分布格局主要为聚集分布。研究表明(宋萍等,2005),集群分布能更好地改变微气候和小生境(如根系分泌物的影响,或利于其生长发育的小气候),比单独个体有较大的抵抗不良环境的能力。因此,集群分布对香果树种群的存在是有利的。不同生境下香果树种群格局动态是从集群分布到随机分布,显示出扩散的趋势。香果树的萌生能力较强,尤其是当母树受到破坏时,将产生大量的萌生苗,使得幼树阶段呈聚集分布;随着个体生态位空间的扩张,种内和种间竞争加剧,种群密度下降,种群的格局转为随机分布。香果树种群不同大小级分布格局的这种动态变化反映了种群在生长发育过程中的一种生态策略和适应机制。这与蔡飞(2000)对青冈(Cyclobalanopsis glauca)种群结构和动态的研究结果相一致。
珍稀物种的保护是保护生物学的一个重要研究领域,它涉及到人类生活的方方面面(Nunes et al., 2001)。目前,香果树正处于濒危灭绝的边缘,亟需加以人工保护。针对香果树始花期长这一生物学特性,首先应采取相应的措施保护母树,尤其是20年生以上的母树;另外,针对自然状态下香果树种子量大而实生苗极为少见的现象,应加强对香果种子雨和种子库,以及幼苗生长动态变化的研究,以找出影响种群更新的主要限制因子,从而为香果树种群的复壮提供依据。
蔡飞. 2000. 杭州西湖山区青冈种群结构和动态的研究. 林业科学, 36(3): 67-72. |
郝朝运, 刘鹏, 徐根娣. 2004. 浙江磐安蕨类植物区系的研究. 浙江师大学报:自然科学版, 27(3): 283-287. |
甘聃, 陈发菊, 梁宏伟, 等. 2006. 珍稀濒危植物香果树种子萌发特性研究. 种子, 25(5): 27-30. |
江洪. 1992. 云杉种群生态学. 北京: 中国林业出版社, 7-139.
|
姬飞腾, 李凤兰, 高述民, 等. 2005. 香果树体细胞胚胎发生. 植物生理学通讯, 41(5): 619-621. |
李钧敏, 金则新. 2006. 珍稀濒危植物香果树ISSR-PCR反应体系的筛选与优化. 安徽农业大学学报, 33(4): 458-461. DOI:10.3969/j.issn.1672-352X.2006.04.004 |
李铁华, 周佑勋, 段小平, 等. 2004. 香果树种子休眠和萌发的生理特性. 中南林学院学报, 24(2): 82-84. DOI:10.3969/j.issn.1673-923X.2004.02.010 |
刘建泉. 2004. 祁连山保护区青海云杉种群分布格局的研究. 西北林学院学报, 19(2): 152-155. DOI:10.3969/j.issn.1001-7461.2004.02.044 |
刘军. 2003. 国家Ⅱ级重点保护植物香果树的保护与利用. 甘肃科技, 19(10): 151-152. DOI:10.3969/j.issn.1000-0952.2003.10.089 |
宋萍, 洪伟, 吴承祯, 等. 2005. 珍稀濒危植物桫椤种群结构与动态研究. 应用生态学报, 16(3): 413-418. DOI:10.3321/j.issn:1001-9332.2005.03.004 |
王昌腾. 2005. 浙江省九龙山国家级自然保护区珍稀濒危植物调查研究. 安徽农业科学, 33(2): 250-251. DOI:10.3969/j.issn.0517-6611.2005.02.037 |
韦小丽, 朱忠荣, 廖明, 等. 2005. 香果树组织培养技术研究. 种子, 24(10): 27-29. |
熊丹, 陈发菊, 李雪萍, 等. 2006. 神农架地区濒危植物香果树的遗传多样性研究. 西北植物学报, 26(6): 1272-1276. DOI:10.3321/j.issn:1000-4025.2006.06.032 |
谢玉芳, 潘林, 杨玉芳, 等. 2004. 香果树育苗技术. 江苏林业科技, 31(2): 39-40. DOI:10.3969/j.issn.1001-7380.2004.02.016 |
徐小玉, 姚崇怀, 潘俊. 2002. 湖北九宫山香果树群落结构特征研究. 西南林学院学报, 22(1): 5-8. |
杨旭, 于明坚, 丁炳扬, 等. 2005. 凤阳山白豆杉种群结构及群落特性的研究. 应用生态学报, 16(7): 1189-1194. DOI:10.3321/j.issn:1001-9332.2005.07.002 |
闫淑君, 洪伟, 吴承祯, 等. 2002. 武夷山天然米槠林优势种群结构与分布格局. 热带亚热带植物学报, 10(1): 15-21. DOI:10.3969/j.issn.1005-3395.2002.01.003 |
于永福. 1999. 中国野生植物保护工作的里程碑——国家重点保护野生植物名录(第一批)出台. 植物杂志, 5: 3-11. |
Arista M. 1995. The structure and dynamics of an Abies pinsapo forest in southern Spain. Forest Ecology and Management, 74: 81-89. DOI:10.1016/0378-1127(94)03507-S |
Gittins R. 1985. Canonnical analysis a review with applications in ecology. Berlin: Springer Verlag.
|
Greig-Smith P. 1952. The use of random and contiguous quadrats in the study of the structure of plant communities. Annual Botany, 16: 293-316. DOI:10.1093/oxfordjournals.aob.a083317 |
Nunes P A L D, Jeroen C J M, Van den Bergh. 2001. Economic valuation of biodiversity: sense or nonsense?. Ecological Economics, 39: 203-222. DOI:10.1016/S0921-8009(01)00233-6 |
Quinn J F, Dunhama A E. 1983. On hypothesis testing in ecology and evolution. American Naturalist: 602-617. |
Wu X P, Zheng Y, Ma K P. 2002. Population distribution and dynamics of Quercus liaotungensis, Fraxinus rhynchophlla and Acer mono in Dongling Mountain, Beijing. Acta Botanica Sinica, 44(2): 212-223. |