 2007, Vol. 43
2007, Vol. 43文章信息
- 卢军, 李凤日.
- Lu Jun, Li Fengri.
- 樟子松人工林的节子寿命及年轮丢失数
- Long and Missing Ring of Knots in Pinus sylvertris var. mongolica Plantation
- 林业科学, 2007, 43(12): 16-21.
- Scientia Silvae Sinicae, 2007, 43(12): 16-21.
-
文章历史
- 收稿日期:2007-07-19
-
作者相关文章



采用较小的初植密度和较短轮伐期是目前人工林经营中的一个趋势(Tappeiner et al., 1986)。但是, 这样会导致树木年轮宽度增大、木材密度减小、尖削度增大和节子数量增加, 导致木材质量降低。枝条是影响木材质量的一个重要因子(Whiteside et al., 1977), 其旺盛生长必然产生大尺寸的节子。在枝条生长过程中, 当枝条基部停止形成年轮时, 枝条并不是立刻死亡(Li et al., 2002), 而会存活几年, 个别枝条会存活10多年(Kershaw et al., 1990; Fujimori, 1993; Mäkinen, 1999)。
树木年轮是树干或枝条横断面上由早材和晚材构成的同心"环带"。但在某些年份, 由于受外界环境条件的制约, 使年轮环带产生不完整的现象, 这就称为年轮变异(ring anomaly)。当变异的年轮数量较多、宽度很小时, 以年轮计年的方法不再可行。常见的年轮变异有伪年轮、断轮和丢失年轮等。在树干基部, 某些年份的年轮很难用肉眼分辨出来, 这种现象称为年轮丢失。在干旱、半干旱地区及森林边缘的树木常见到年轮消失的现象。引起年轮消失的原因较复杂, 一种情况是有的年份树木基部形成层没有完全分化, 即根本没有形成年轮; 另一种情况是有的年份只分化出相当于晚材的那部分年轮, 由于这种年轮中的细胞大小、细胞壁厚度和木材颜色同晚材接近, 使之与上1年晚材合在一起, 不易分辨。这样, 就不易测得该树木的真实年龄, 当然测得的某一断面的年轮宽度也必然不能反映树木的实际生长量。枝条年轮消失现象多产生在树冠下部的枝条, 而树干上的年轮消失则多出现在树冠以下部分, 其原因是枝条或树冠所生产的有机物不能满足枝冠自身的消耗, 因而, 对主枝或树干的贡献很少或没有(Jia et al., 2005)。结合树木的生长过程研究枝条丢失年轮数及枝条的寿命可以了解枝条在发育过程中生长、受抑制和死亡的信息。戚大伟等(2001)应用计算机数字图象处理技术对木材的年轮进行了检测, 采用区域生长技术填补了某些丢失的年轮, 应用模式识别技术对年轮图像自动识别提供了切实可行的研究手段。
本文利用樟子松(Pinus sylvestris var. mongolica)枝条解析和节子剖析数据, 分析节子丢失年轮数及其频数分布的变化规律, 研究胸径、冠幅等因子对平均节子寿命及丢失年轮数的影响, 为樟子松人工林的抚育间伐和人工整枝提供依据。
1 试验区概况试验地位于黑龙江省佳木斯市桦川县林业局横头山林场和佳木斯市孟家岗林场。横头山林场位于桦川县西南部(130°28'29″-130°44'14″E, 40°34'40″-46°34'14″N), 平均海拔350 m。植被属完达山植物区系(据中国森林立地分类系统划分), 现有林相以人工林为主天然次生林为辅。大陆性季风气候, 年均气温2.5℃, 极端最高气温36℃, 最低气温-39℃, 年≥ 10℃有效积温2 538℃, 全年日照时数2 542 h, 年降水量约460 mm, 全年无霜期130 d左右。孟家岗林场位于桦南县东北部(130°32'42″-130°52'36″E, 46°20'-46°30'50″ N), 距县城21 km。平均海拔250 m。植被属小兴安岭-张广才岭亚区。大陆性季风气候, 年均气温2.7℃。极端最高气温35.6℃, 最低气温-34.7℃, 年≥ 10℃有效积温2 547℃。年平均降水量550 mm, 全年日照时数1 955 h, 无霜期120 d左右。
2 研究方法2004-2006年, 在黑龙江省佳木斯市孟家岗林场和横头山林场, 选择不同年龄、不同立地和不同密度的樟子松人工林共设置了14块标准地, 各标准地面积为0.04~0.20 hm2; 林分年龄18~47年; 林分密度为346~4 240株·hm-2; 郁闭度均大于0.6。在每个标准地中采用等断面径级标准木法选取了5~7株标准木。
2.1 节子年龄测定1) 伐倒解析木后, 按照1 m的区分段, 把树干部分锯开, 测量每个死枝距离地面的高度, 并测定每个死枝的方位角。节子的位置根据枝条死亡后在树干上的疤痕来确定。
2) 在树冠基部以下, 截取20~35 cm长的木段带回实验室, 并对每轮的死枝进行编号。所截取的木段的数量与树干的长度相关, 一般每株解析木可获得12~16段, 每段上都包含1轮死枝。
3) 每个木段的一轮死枝约有5~8个死枝, 受剖解方法限制, 只选取两个最大的死枝, 两个死枝之间的夹角为30°~60°, 沿树干方向用手持油锯通过树干髓心进行纵剖, 得到2个死枝的纵剖面, 图 1是其中的1个剖面。

|
图 1 节子纵向剖析图 Fig. 1 Chart of knot dissected in longitudinal direction |
4) 把获取的所有节子剖面带回实验室, 用扫描仪进行扫描, 采用年轮图像分析系统(WinDENDROTM V6.5), 测定节子的年轮数, 包括以下4部分:1)髓心到枝条停止形成年轮处的年轮数(RingPC); 2)从髓心到死枝开始变疏松(即死亡)处的年轮数(RingPL); 3)从髓心到节子被完全包藏时的年轮数(RingPW); 4)从髓心到树皮部的年轮数(RingPB)。通过图像计年得到枝条的寿命(long)和枝条的观测年轮(ring)。
2.2 节子数据分析从图 1中可以看出从髓心到死枝开始变疏松(即死亡)处(图 1中黑色的部位)的年轮数就是节子寿命。
而RingPC就是可观测的枝条的年轮数Ring。枝条停止生长到彻底死亡之间所形成的丢失年轮数则是节子的寿命减去髓心到枝条停止形成年轮处的年轮数。
有的节子并没有达到完全包藏阶段, 有的节子只是处于停止生长阶段, 因此, 这种节子不存在丢失年轮的现象, 在数据处理过程, 这些节子的数据被剔除。所有的节子图像年轮纪年所获得的数据用来计算丢失年轮数在节子寿命期中所占的比例和丢失年轮数。在实际的图像计年过程中, 由于图像本身和人眼分辨力的限制, 可能仍会有非常细小的年轮形成了但却无法观测出来。
本研究共收集标准样木71株, 剖析所得节子数据2 352个, 经过数据的预处理, 剔除其中的错误数据(主要是由于剖析过程中出现的异常节子)和未形成疏松节的(不可确定丢失年轮数), 最终用于分析的节子数据为2 126个。各标准木的基本测树因子见表 1。全部2 126个节子的寿命、年轮数、丢失年轮数和丢失年轮比例的统计量见表 2。
|
|

|
|
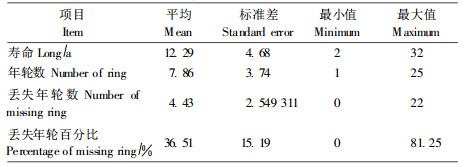
樟子松人工林节子丢失年轮0~22个, 其均值为4.4个, 丢失年轮数所占的比例平均为36.5%(表 2)。
由图 2可知, 樟子松人工林节子发生丢失年轮是普遍现象, 绝大多数(占99.8%)的节子会出现丢失年轮现象, 仅仅0.2%的节子未丢失年轮。73%的节子丢失了5个和5个以下的年轮数, 但丢失1年及以下的节子占5.6%。有83.2%的节子丢失了2~7个年轮, 88.6%的节子丢失了1~7个年轮, 丢失的年轮数大于7的节子所占的比例仅为11.2%。

|
图 2 年轮丢失相对频率和累积频率 Fig. 2 Relative and cumulative frequency of the number of missing rings |
为了反映樟子松人工林节子丢失年轮数与树龄的关系, 根据各标准木的年龄计算了不同龄阶樟子松节子丢失年轮的平均数(图 3)。由图 3可知, 年龄为20年以下和20~30年间的樟子松人工林的节子年轮丢失现象并不是很明显, 平均小于50个; 但随着年龄的增大, 丢失年轮数急剧增长。在30~40年的樟子松人工林中, 平均丢失个数增至130左右, 当年龄超过40年以上时, 平均丢失个数超过了170。

|
图 3 年轮丢失数量与年龄的关系 Fig. 3 Retation between total missing ring and tree age |
樟子松人工林丢失年轮的节子数量随着节子的着生高度呈规律性的变化(表 3)。全部用于分析的2 126个节子中, 丢失年轮的节子个数在树干4 m以下最多(占55.7%), 都保持在200个以上。随着高度的增加, 丢失年轮的节子数呈明显减少的趋势, 其变化趋势明显。节子的寿命和节子可测年轮数在树干基部(0~7 m)随着节子高度的增加而增大, 之后(>7 m)呈稳定状态。节子丢失年轮数在树干基部(0~6 m)随节子高度的增加缓慢增加, 之后呈平稳状态, 当节子高度大于9 m后略有下降。
|
|
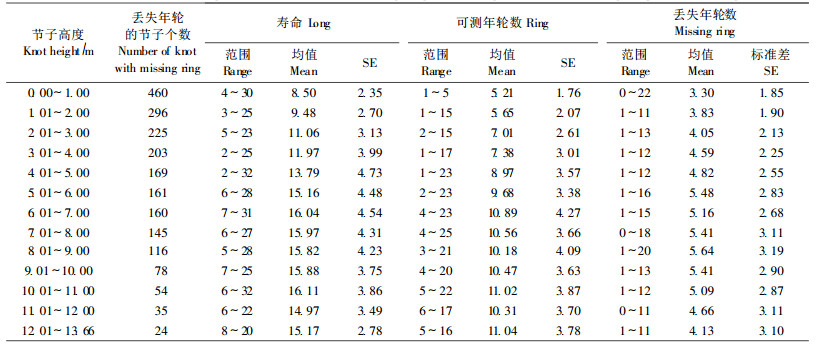
樟子松人工林节子的可测年轮数与节子的寿命成正比, 是明显的线性递增关系(图 4), 丢失年轮所占比例的百分比与节子的寿命之间没有很明显的递减关系, 只是在总体上呈现一种减少的趋势(图 5)。节子的寿命越长, 则节子丢失年轮数量就越多, 它们之间呈线性递增关系, 如图 6所示。

|
图 4 节子年轮数与寿命的关系 Fig. 4 Relationship between knot ring and long |

|
图 5 节子丢失年轮数百分比与寿命的关系 Fig. 5 Relationship between percentage of knot missing ring and long |

|
图 6 丢失年轮数与节子寿命的关系 Fig. 6 Relationship between number of missing ring and long |
所研究的14块标准地的立地条件和林分密度均不相同, 因此樟子松的生长状况也不尽相同, 各标准地内樟子松的胸径生长有较大的差异。分析胸径与每株标准木节子平均丢失年轮数的情况, 发现平均丢失年轮数随着树木胸径的增大而呈增长的趋势(图 7), 胸径6~15 cm时, 平均丢失年轮数3个左右, 胸径20 cm左右时, 平均丢失4~5个年轮, 胸径>25 cm时, 平均丢失年轮6~8个, 如图 7所示。

|
图 7 节子平均丢失年轮数与胸径的关系 Fig. 7 Relationship between mean number of missing ring and DBH |
树木胸径的生长促使枝条分化, 树木出现自然整枝, 死枝逐渐形成, 并且随着胸径的不断生长节子被树干的木质部所包藏。胸径与死枝(节子)的寿命之间是相关的, 樟子松人工林林木的胸径越大, 节子的存活时间就会越长, 节子的寿命也会越大, 呈明显的线性递增关系(图 8)。

|
图 8 节子的寿命与树木胸径的关系图 Fig. 8 Relationship between long of knot and DBH |
树冠的大小直接影响活枝的光合作用, 大的树冠显示出树木强的生活力, 其枝条不容易死亡, 可以维持较长的生活期, 随着枝条死亡, 丢失年轮数增加; 而小的树冠处于压制状态, 枝条容易死亡, 则丢失的年轮数会较少。用冠幅来描述树冠的活力, 分析樟子松人工林冠幅与节子丢失年轮数之间关系, 结果表明节子的丢失年轮数与冠幅成反比, 即随着冠幅增加节子的丢失年轮数逐渐下降(图 9)。由图 9可知, 樟子松节子丢失的年轮集中分布于冠幅为1.0~3.0 m的树木上, 丢失数量为2~8个。

|
图 9 节子丢失年轮数与冠幅的关系图 Fig. 9 Relationship between number of missing ring of knot and crown width |
本研究中, 死枝不形成可观测的年轮的数量为0~22年, 其均值为4.4个, 丢失年轮数所占的比例平均为36.5%, 这说明在枝条的生长过程中, 有1 3的时期处于对树干生长无贡献状态, 甚至会阻碍树干木质部的生长。因此, 枝条形成丢失年轮的时期即为整枝的最佳时期。接近60%的节子丢失年轮数≤ 4个, 64.72%的节子丢失年轮2~7个, 7.63%的节子丢失的年轮数大于7。这些结果与以往的活枝丢失年轮的研究结果相类似, Reukem (1959)研究表明, 美国西部花旗松活枝基径一般有9~10个不可观测年轮; Andrews等(1939)对美国黄松(Pinus ponderosa)的研究表明, 活枝最多产生34个不可观测年轮, 且有50%的枝条的丢失年轮数超过了10个。
本研究认为年龄会对丢失年轮数产生影响。20年以下和20~30年间的樟子松人工林的节子年轮丢失现象并不是很明显, 随着年龄的增大, 丢失年轮数急剧增长。
节子在树干中的着生位置决定了节子被包藏可能性, 在树干下部的位置, 节子几乎都是被完全包藏, 这主要是由于幼树阶段的樟子松所形成的细小枝条死亡后, 被逐渐增长的树干木质部所包藏; 而树干中上部形成的枝条一般属于较大的枝条, 自然整枝刚停止或者停止几年, 节子刚死亡或死亡若干年, 不容易被完全包藏。樟子松丢失的年轮个数在树干的4 m处以下是最多的, 随着高度增加, 丢失年轮数呈明显的减少趋势。
节子的可测年轮数与节子的寿命成正比, 呈线性递增关系, 但是丢失年轮数所占百分比与节子寿命之间没有很显著的递减关系, 只是在总体上呈一种减少的趋势。随着节子寿命增加, 节子丢失年轮数增加, 但它们之间不是显著的线性关系, 而是逐渐递增的。
戚大伟. 2001. 木材无损检测图像处理系统的研究. 林业科学, 37(16): 92-96. |
Andrews S R, Gill L S. 1939. Determining the time branches on living trees have been dead. Journal of Forest, 37: 930-935. |
Celine M, Yves C. 2002. External indicators of living branches with missing rings within a tree crown of Corsican pine. Forestry, 75(5): 569-578. DOI:10.1093/forestry/75.5.569 |
Craig G L, Sally E D. 1999. Frequency of partial and missing rings in Acer saccharum in relation to canopy position and growth rate. Plant Ecology, 143: 189-202. DOI:10.1023/A:1009847819158 |
Fritts Harold C, Fujimori T. 1993. Dynamics of crown structure and stem growth based on knot analysis of a hinoki cyspress. Forest Ecology and Management, 56: 57-68. DOI:10.1016/0378-1127(93)90103-T |
Jia W W, Li F R. 2005. Distribution, size and development phases of Knots for Pinus sylvestris L. var. mongolica Litvin in northeast China. Joural of Korean Forestry Society :English Version, 94(5): 313-320. |
Kershaw JA, Maguire D A. 1990. Longevity and duration of radial growth in Douglas-fir branches. Canadian Journal of Forest Research, 20: 1690-1695. DOI:10.1139/x90-225 |
Li F R, Hee Moon Yang. 2002. Diameter and length growth of primary branches in a Larix olgensis plantation. Journal of Korean Forest Society, 91(2): 219-230. |
Mäkinen H, Colin F. 1998. Predicting branch and angle and branch diameter of Scots pine from usual tree measurements and stand structure information. Canadian Journal of Forest Research, 28: 1686-1696. DOI:10.1139/x98-141 |
Mäkinen H. 1999a. Growth, suppression, death, and self-pruning of branches of Scots pine in southern and central Finland. Canadian Journal of Forest Research, 29: 585-594. DOI:10.1139/x99-026 |
Mäkinen H. 1999b. Effect of stand density on radial growth of branches of Scots pine in southern and central Finland. Canadian Journal of Forest Research, 29: 1216-1224. DOI:10.1139/x99-060 |
Reukema D L. 1959. M issing annual rings in branches of young growth Douglas-fir. Ecology, 40: 480-482. DOI:10.2307/1929767 |
Tappeiner J C, Knapp W H, Wiermann C A. 1986. Silviculture, the past 30 years, the next 30 years.Part Ⅱ.The Pacific Coast. Journal of Forest, 84: 37-46. |
Whiteside I D, Wilcox M D, Justin J R. 1977. New Zealand Douglas-fir timber quality in relation to silviculture. New Zealand Journal of Forest, 22: 24-45. |