 2007, Vol. 43
2007, Vol. 43文章信息
- 郑维列, 薛会英, 罗大庆, 王景升, 罗建.
- Zheng Weilie, Xue Huiying, Luo Daqing, Wang Jingsheng, Luo Jian.
- 巨柏种群的生态地理分布与群落学特征
- Eco-geographic Distribution and Coenology Characteristics of Cupressus gigantea
- 林业科学, 2007, 43(12): 8-15.
- Scientia Silvae Sinicae, 2007, 43(12): 8-15.
-
文章历史
- 收稿日期:2006-09-15
-
作者相关文章



2. 西藏农牧学院 林芝 860000;
3. 西藏高原生态研究所 林芝 860000
2. Tibet Agriculture and Animal Husbandry University Nyingchi 860000;
3. Research Institute of Tibet Plateau Ecology Nyingchi 860000
巨柏(Cupressus gigantea)为柏科(Cupressaceae)柏木属(Cupressus)常绿乔木, 属西藏特有树种(郑万钧等, 1975; 1978;吴征镒, 1983)。多数学者认为它可能是柏科植物中残遗的原始类型(中国科学院青藏高原综合科学考察队, 1988; 梁红平, 1990)。据文献记载, 巨柏仅分布在雅鲁藏布江中游朗县至米林、林芝一带的河谷山地。由于自然历史因素和现代人类活动的影响, 其个体数量日益减少, 分布地域逐渐狭窄、孤立, 处于濒危状态, 被列为国家重点保护植物(于勇福, 1999; 汪松等, 2004)。
从发现巨柏至今已有30多年的历史, 但由于分布地理位置独特或其他的一些特殊原因, 对它的研究仍不够深入。已有的研究仅限于其生物学特性、分布、生长、引种育苗技术以及保护现状等方面(黄健等, 1993; 胡匡祜等, 1993; 陈端, 1994; 1995;1996;李乾振等, 2003; 王景升等, 2005; 吴兴等, 2005)。对于巨柏生境特点、群落特征及种群现状等方面的研究未见报道, 其濒危原因和机理仍不是很清楚。鉴于目前的研究现状和巨柏这一古老树种对西藏及东喜马拉雅地区植物区系的演化历史、古生物学、古气候学等方面研究的重要学术价值, 本文在调查的基础上, 结合相关文献, 对巨柏的生态地理分布现状、群落植物区系特点、群落学特征和种群结构等方面作了分析与研究, 目的是为巨柏这一濒危物种的深入研究和有效保护提供基础资料和科学依据。
1 研究区概况研究区位于西藏自治区东南部, 雅鲁藏布江中下游流域及其支流尼洋河、易贡藏布的部分地段。地形为典型的高山峡谷, 包括东喜马拉雅山脉北侧和念青唐古拉山脉以南部分山地。该区承接印度洋季风北进的作用和减弱影响, 加之高山、峡谷的巨大高差, 水平和垂直地带的温度、降水差异较大, 气候类型多样。由东向西依次为亚热带山地季风湿润和半湿润气候区、高原温带季风半湿润气候区和高原温带季风半干旱气候区(中国科学院青藏高原综合科学考察队, 1984)。研究区年均降水量400~1 100 mm, 年均温度7~12 ℃, 年均日照时数1 500~2 600 h, 全年无霜期180~320 d以上。山体主要由花岗岩和变质岩构成, 土壤以棕色森林土为主。
由于水平和垂直地带的气候差异, 在本区山地不同垂直带上, 几乎集中了从热带到极地寒带的各种植被类型。研究区海拔2 200~2 800 m为亚热带常绿阔叶林和常绿针阔叶混交林、海拔2 800~3 500 m为温性针阔叶混交林。植被建群种主要有松科(Pinaceae)的冷杉属(Abies)、云杉属(Picea)和松属(Pinus), 柏科(Cupressaceae)柏木属(Cupreaaus)、圆柏属(Sabina), 杜鹃花科(Ericaceae)杜鹃花属(Rhododendron)以及壳斗科(Fagaceae)栎属(Quercus)植物。植被区系属泛北极植物区中国-喜马拉雅植物亚区东喜玛拉雅地区(李文华, 1985)。
2 研究方法 2.1 地理分布对巨柏分布区进行踏查, 用GPS仪(误差 < 15 m)对巨柏的水平和垂直分布地域进行实地测量和边界定位, 确定其地理分布区, 并在1:10万的地图上勾绘标注; 种群分布区的确定主要根据各种群间断分布的距离, 同时结合气象资料和相关文献分析生态类型差异(李文华, 1985), 然后进行划分, 绘制成地理分布图。
2.2 群落调查在种群划分的基础上, 在各种群分布区内选择典型地段, 分别设置调查样地, 共设20 m×30 m的样地12块, 其中林芝种群3块样地, 米林种群3块样地, 朗县种群6块样地。样地内调查内容包括乔木树种的每木检尺, 实测其胸径、树高; 每个样地按对角线法设置2 m×2 m的灌木样方和1 m×1 m的草本小样方各5个, 样方内灌木和草本植物分种计录个体数量、高度和盖度, 同时记录灌木样方中高度 < 1.5 m的乔木树种的幼树、幼苗的数量、高度及地径。将所收集数据进行群落学特征分析。
2.3 区系组成与地理成分分析将所有样地采集的植物标本带回实验室进行鉴定, 并结合西藏高原生态研究所标本室历年采集的标本, 分析其区系组成。按照吴征镒先生对中国种子植物分布区类型的划分方法(吴征镒, 1991), 将巨柏群落调查到的种子植物合并进行地理成分分析。
2.4 群落外貌群落中各物种的生活型以及叶的性质按照Raunkiaer的分类系统进行划分(Raunkiaer, 1934; 武吉华, 1995), 并绘制生活型图谱进行分析。
2.5 种群结构年龄结构是种群的重要特征之一, 能够客观地反映种群发展的动态变化趋势。由于巨柏立木胸径粗大, 无法完整获取其年龄数据, 鉴于多数乔木树种的胸径与年龄具有较好的相关性, 对巨柏种群结构我们采取高度加径级结构替代年龄结构的方法(Knowles et al., 1983; Parker et al., 1984; 梁士楚, 1992; 蔡飞等, 1997), 将巨柏种群按胸径级大小划分成9个径级。对巨柏种群径级的划分按下述2种方式:1)对于胸径小于2.5 cm、高度不足1.5 m的幼树和幼苗, 统一按高度进行分级:Ⅰ级的高度1.0~30.0 cm, Ⅱ级的高度30.0~60.0 cm, Ⅲ级的高度60.0~150.0 cm; 2)胸径2.5 cm以上的幼树和大树则按整化的胸径级进行划分, 以50.0 cm为径阶划分出5个径级, Ⅳ级为2.5~55.5 cm、Ⅴ级为55.5~105.5 cm、Ⅵ级为105.5~155.5 cm、Ⅶ级为155.5~205.5 cm、Ⅷ级为155.5~205.5 cm, Ⅸ级为胸径255.0 cm以上的全部巨树。按上述方法对不同巨柏种群进行统计和划分后, 绘制成种群结构图。
3 结果与分析 3.1 巨柏生态地理分布特征图 1所示, 巨柏天然分布于雅鲁藏布江中下游河谷及其支流尼洋曲、易贡藏布的沿江地段, 该区域位于西藏东南部, 行政隶属林芝地区。巨柏分布区大致呈西南——东北走向, 从朗县开始, 沿雅鲁藏布江经米林县、林芝县, 最远至雅鲁藏布江大拐弯顶部的易贡湖区, 流域全长约430 km。水平地理分布范围为29°02′—30°16′ N, 93°03′—94°54′ E, 分布海拔为2 250~3 550 m。巨柏主要沿雅鲁藏布江河谷两岸呈狭带状、间断式分布, 形成了彼此间隔、大小不等的4个种群, 种群面积由西向东逐渐缩小。分布区受季风的影响, 水、热变化较大, 降雨量由东向西递减, 干燥度增加, 在水平地带上跨越了湿润、半湿润区和半干旱区3大生态区, 各种群的生态环境差异较大。根据各种群间断分布距离和生态环境的差异, 将其划分为4个种群进行分析和研究。

|
图 1 巨柏种群地理分布图 Fig. 1 Geography distribution of C.gigantea populations |
朗县种群位于朗县与里龙之间(29°02′58″—29°08′45″ N, 93°03′30″—93°51′04″ E), 海拔2 900~4 500 m, 该区地形狭窄, 山势陡峭, 土质沙化, 属于藏东南森林向灌丛、草原过渡区, 河谷地带森林基本消失, 只在海拔3 500~4 200 m外的山地中上部还有小面积森林分布。河谷区环境干燥, 年均温8.9 ℃, 年降水量420 mm左右, 年蒸发量2 682 mm, 是降水量的6.3倍, 植被以旱生灌丛为主, 巨柏群落在该区域的出现, 形成了半干旱河谷地带的特殊植被景观。巨柏主要在沿江两岸海拔2 960~3 550 m的山地中下部分布, 一方面沿江边的夏季最高水位线在砾石河滩上成带状生长, 另一方面在河谷两侧的阳坡和半阳坡沙地、石灰岩出露的坡地或陡壁的裸露岩石缝隙中生长并形成疏林。朗县种群是分布区中最大的一个种群, 其中, 朗县与甲格之间巨柏呈连续分布, 种群密度较大, 为438株·hm-2。
3.1.2 米林种群米林种群位于朗县下游170 km处(29°20′24″—29°24′21″ N, 94°22′40″—94°25′20″ E)。该区属雅鲁藏布江宽阔河谷半湿润森林区, 年均气温8.2 ℃, 年降水量700 mm左右, 全年降水量的69%集中在夏季(6—9月), 冬春季节为干旱季, 年蒸发量1 201 mm。河谷地带风沙地貌发育明显, 沙地呈斑块状分布于河心滩、河漫滩、河谷阶地和局部谷坡上; 河谷两侧山地生境较湿润, 植被丰富, 海拔3 500 m以上分布着急尖长苞冷杉(Abies georgei var. smithii)林, 其下是高山松(Pinus densata)林, 山体下部多灌丛和灌状川滇高山栎(Quercus aquifolioides)林。巨柏集中分布于雅鲁藏布江北岸、在海拔2 920~3 100 m的江边石滩地以及阳坡中下部高山松林与灌丛之间, 呈斑块状或带状间断分布, 种群密度155株·hm-2。
3.1.3 林芝种群林芝种群位于雅鲁藏布江一级支流尼洋曲流域的八一镇附近(29°37′49″—29°37′32″ N, 94°23′27″—94°24′32″ E)。该区亦属宽阔河谷半湿润森林区, 年均气温8.7 ℃, 年降水量688 mm, 年蒸发量1 792 mm。巨柏集中分布于尼洋曲东岸巴结乡附近海拔3 040~3 250 m的坡地上, 占地面积约10.6 hm2, 处于高山栎灌丛的下部, 生境是沙质土的半阳坡。虽然该种群的分布面积小于朗县和米林种群, 但巨柏的生长状况明显优于它们, 表现为巨柏树体粗壮高大并形成郁闭的林分, 平均密度87株·hm-2, 90%的单株平均年龄在千年以上。由于该处巨柏林是当地村庄附近的风水林所以才得以保护至今, 目前已开发为当地的特殊景观和旅游风景保护区。
3.1.4 易贡种群易贡藏布流域的易贡湖区处于藏东南亚热带的北缘, 属山地温暖湿润季风气候区。由于承印度洋暖湿气流沿雅鲁藏布江河谷北进的作用, 该区雨量充沛, 气候温和润湿, 年均气温11.4 ℃, 年降水量960~1 100 mm, 降雨集中在5—9月, 占全年降水量的78%;年蒸发量1 458 mm。本区属于亚热带常绿阔叶林区, 森林面积大, 树种多, 植被类型复杂而多样, 在海拔2 700 m以下, 为通麦栎(Quercus tungmeiensis)等组成的硬叶常绿阔叶林和常绿阔叶林、落叶阔叶林, 其间常有西藏柏木(Cupressus sorulosa)、云南铁杉(Tsuga dumosa)和华山松(Pinus armandi)林分布, 其他树种有云南松(Pinus yunnanensis)、乔松(Pinus griffithii)、云南红豆杉(Taxus wallichiana)、桦木(Betula spp.)等; 在海拔2 700 m以上是分布面积最大、垂直带幅较宽的云、冷杉林分布地带。通过调查发现, 该区域分布的巨柏仅剩几株孤立木(典型株树龄1 600年左右, 胸径83.4 cm, 树高41.8 m), 分布在海拔2 250 m的湖畔平缓坡麓农田或村落旁, 其下未见幼苗、幼树。鉴于该分布区与其他巨柏分布区的距离较远, 我们认为该种群处于孤立的特殊状态, 因此未将其纳入本文的种群分析。
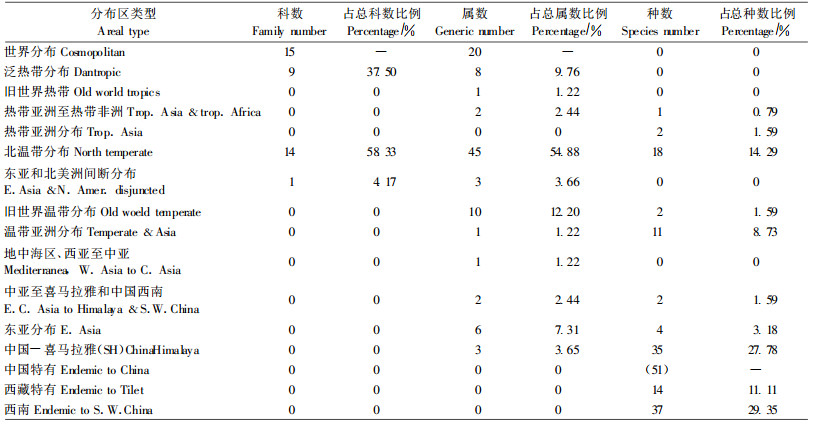
3.2 巨柏群落区系组成与地理成分根据3个巨柏群落, 12个样地的调查, 群落中共有维管束植物130种, 分别隶属43科, 106属。其中蕨类植物4科, 4属, 4种; 裸子植物2科, 2属, 2种; 被子植物37科, 100属, 124种(其中单子叶植物4科, 15属, 21种; 双子叶植物33科, 85属, 103种)。群落植物含种数最多的是菊科(Compositae) 14种, 其次是禾本科(Gramineae) 13种、蔷薇科(Rosaceae) 12种、唇形科(Labiatae) 10种、豆科(Leguminosae) 7种、伞形科(Umberlliferae) 6种、毛茛科(Ranunculaceae) 5种、石竹科(Caryophyllaceae) 4种、十字花科(Cruciferae) 4种、鼠李科(Rhamnaceae) 4种、龙胆科(Gentianaceae) 4种、莎草科(Cyperaceae) 4种、紫草科(Boraginaceae) 3种、玄参科(Scrophulariaceae) 3种、百合科(Liliaceae) 3种, 以上15科占总科数的35%;其余组成中含2种的有6科, 占总科数的14%;仅含1种的有22科, 占科总数的51%。在属的组成中仅含1种的有82属, 占总属数的77%。
由以上分析可知, 巨柏群落植物的科、属组成分散, 与种的数目相比, 科、属数目相对较多, 尤其是属的组成最突出, 这一现象说明巨柏群落属于一个比较古老的植物区系。
将巨柏群落中调查到的39科102属126种种子植物合并进行地理成分分析(表 1)。在科的分布类型中, 北温带分布的有14科, 占总科数的58.33%;其次为泛热带分布, 有9科, 占总科数的37.50%。在102属植物中, 温带性质的属有59个, 占总属数71.96%, 其中主要以北温带分布及其变型的属为主, 共有45属, 占总属数的54.88%, 旧世界温带分布的属有10个, 占总属数的12.20%, 群落中热带性质的属较少, 共11属, 占总属数的13.42%, 其中以泛热带分布的属种多, 8属, 占总属数的9.76%, 其余分布类型的比例较小。扣除世界分布属, 属的分布类型表明巨柏群落具有明显的温带性质。从种的分布类型来看, 以中国特有分布的成分最多, 有51种, 其中, 西南分布的有37种, 占总数的29.35%, 西藏特有种14种, 占总数的11.11%。其次为中国-喜马拉雅分布种, 有35种, 占总数的27.78%;再次是北温带和温带亚洲分布的种, 分别有18、11个种, 占总数的14.29%、8.73%, 其余分布类型的比例较小。种的组成成分中含有大量的中国(西藏)特有种和中国-喜马拉雅分布种。
|
|
群落外貌是群落与外界环境条件, 特别是气候条件长期适应的结果(曲仲湘等, 1983)。反映群落外貌最主要的标志是植物的生活型谱、叶的性质等(Daubenmire, 1947)。分析巨柏群落的外貌特征, 有助于进一步认识该区域植被与气候的关系。
3.3.1 生活型由图 2所示, 巨柏群落的地面芽植物占57.69%;高位芽植物占23.08%;1年生植物占9.23%;而地上芽植物和地下芽植物较少。在高位芽植物中以小高位芽和矮高位芽的小型灌木植物居多, 缺少大灌木高位芽植物, 大高位芽植物只有巨柏和高山松2种常绿大乔木。巨柏群落高位芽植物比例表明该区域在生长季节具备一定的水、热条件, 能够满足一些乔木和矮灌木植物的生长需求; 而群落中地面芽植物居多表明该区域还有一个漫长的寒冷、干燥季节。群落生活型基本反映出该区域由温性森林植被向高寒、干旱灌丛、草原植被过渡的特点。

|
图 2 巨柏群落植物生活型谱 Fig. 2 Life form spectrum of plants in C.gigantea communities |
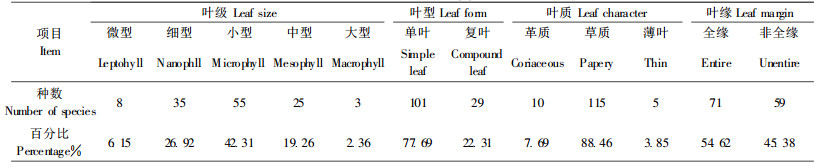
按照Raunkiaer的分类系统, 将巨柏群落中维管束植物叶的性质按叶级、叶型、叶质和叶缘4个方面进行统计分析(表 2)。结果表明, 群落植物的叶级以小型叶为主, 有55种, 占42.31%;其次为细型叶35种, 占26.92%;中型叶25种, 占19.26%;大型叶和微型叶分别为3种和8种, 各占2.36%和6.15%。叶型以单叶为主, 共计101种, 占77.69%, 其余为复叶, 共29种, 占22.31%;叶质中草质叶居多, 共115种, 占88.46%。可以看出, 巨柏群落主要以全缘的草质小型单叶植物为主, 具有明显的落叶性质。
|
|
巨柏群落一般可划分为乔木层、灌木层和草本层, 而层间植物发育较差。巨柏群落通常以巨柏为建群种, 乔木层其他树种高山松、川滇高山栎和光核桃(Prunus mira)多分布于巨柏林缘, 或群落交错区。从各群落的巨柏生长状况来看(表 3), 林芝种群巨柏的平均树高为29.1 m, 平均胸径为139.8 cm, 平均盖度为40%, 生长状况在各种群中最优; 米林种群巨柏的平均树高为12.9 m, 平均胸径为87.1 cm, 平均盖度为10%;朗县种群巨柏的平均树高为6.9 m, 平均胸径为50.7 cm, 平均盖度为30%, 朗县种群的密度在各种群中最大, 平均盖度略大于米林, 但生长状况在各种群中最差。通过上述比较, 各巨柏种群的生长状况排序为林芝种群 > 米林种群 > 朗县种群, 从分布区类型来看, 巨柏的生长状况总体上由半湿润向半干旱分布区逐渐减弱。
|
|
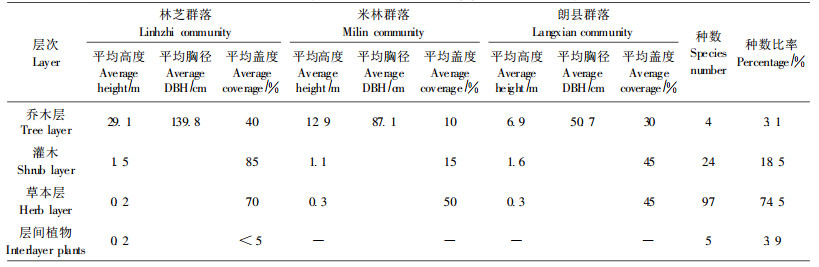
群落中灌木层植物有24种, 主要优势种有美花山蚂蝗(Desmodium elegans)、砂生槐(Sophora moorcroftiana)、西南野丁香(Leptodermis purdomii)、绢毛蔷薇(Rosa sericea)、楔叶绣线菊(Spiraea canescens)、毛叶绣线菊(Spiraea mollifolia)、小叶栒子(Cotoneaster microphyllus)、黑果小檗(Berberis ignorata)、二色锦鸡儿(Caragana bicolor)、川藏香茶菜(Rabdosia pseudo-irrorata)、架棚(Ceratostigma minus)、日喀则蒿(Artemisia xigazeensis)、毛莲蒿(Artemisia vestita)等。灌木层的高度为1.1~1.6 m, 各群落盖度15%~80%。从半湿润区向半干旱区, 巨柏群落灌木层组成种类逐渐减少, 盖度降低。
巨柏群落草本层植物种类最多, 是群落中较发达的层次。共有草本植物97种, 主要由禾本科(Cramineae)、莎草科(Cyperaceae)、菊科(Compositae)、桔梗科(Campanulaceae)等科的植物组成, 各群落的草本层盖度为45%~70%。
3.5 种群结构特征种群结构分析能较好地反映出巨柏各种群的现状和发展趋势(图 3)。

|
图 3 巨柏种群径级结构图 Fig. 3 Diameter class structure of C.gigantea populations |
林芝种群的径级分布连续, 结构基本呈纺锤形。纺锤形结构中部的第Ⅴ级比率最大, 为41.4%, 逐级往上比率依次递减, 结构呈金字塔形, 种群中成年个体占有较大比重, 种群处于成熟阶段, 而Ⅴ级以上的各级比率之和达49.2%, 高出第Ⅴ级的比率, 这说明种群老龄个体的比重也大, 老龄化发展的趋势明显; 从Ⅴ级往下, 第Ⅳ级开始径级比率急剧缩小到7.2%, 并逐级锐减到Ⅰ、Ⅱ级的0.5%, 结构呈倒金字塔形, 种群中的幼龄个体数量极少, 尤其是幼树和幼苗的稀缺, 说明种群的更新不良。可见, 林芝种群正呈现出老龄化的发展趋势, 更新受阻导致种群结构严重失调, 势必会加剧这种趋势。
米林种群的径级分布不连续, 结构不规则。径级结构中Ⅰ径级所占比率较大, 为58.6%, 第Ⅱ和Ⅲ级出现断层; Ⅳ~Ⅶ级所占比率较少, 在6.9%~13.8%之间; Ⅷ级以上没有分布。这种结构表明, 该种群虽然具有较强的繁殖能力, 能产生大量的幼苗和幼株, 但在向幼树过渡的过程中全部死亡, 导致更新过程被阻断, 种群出现严重的断代现象。如此以来, 种群一方面是上层成年个体和老龄大树的不断衰老、死亡, 种群数量不断减少, 另一方由于更新受阻致使种群繁衍扩大困难, 在2方面压力下, 该种群逐渐萎缩成为必然趋势。
朗县种群的径级结构不规则但分布基本连续, 径级结构大致呈金字塔型。基部的Ⅰ径级所占比率最大, 为67.8%;Ⅱ~Ⅶ级各级所占的比率较小, 在1.8%~15.8%之间; 与米林种群相同, 径级顶部Ⅷ级以上没有分布。从朗县种群的径级结构分析来看, 其更新的延续性要比林芝和米林种群好, 种群能够产生大量的幼苗和幼株, 并且维持一定数量的个体向幼树和大树阶段过渡, 保证了种群的连续, 种群基本处于稳定状态, 如果没有较大的干扰影响, 适当采取人为保护和管理措施, 该种群仍将在相当长的时间内延续。
4 结论与讨论巨柏是西藏分布的特有树种, 也是我国柏科植物中残遗的古老树种之一, 其现代地理分布区域狭窄, 仅在雅鲁藏布江中下游流域的朗县与易贡之间的沿江两岸部分河谷地带分布。巨柏的分布方式呈狭带状或斑块状间断分布, 由此形成了大小不等、彼此间隔的4个种群。分布区的水平地理分布范围为29°02′~30°16′ N, 93°03′~94°54′ E, 垂直分布海拔2 250~3 550 m, 虽然分布区范围不大, 但水平地带由西向东跨越了藏东南的半干旱、半湿润和湿润3大生态区。朗县种群位于半干旱区河谷地带, 该区生境干燥、植被稀疏, 但巨柏在该区能够连续分布, 种群密度和分布范围居所有种群之首, 是分布区中最大的一个种群; 林芝和米林种群位于半湿润区, 其分布范围和种群密度远小于朗县种群, 属小种群; 易贡种群处于环境最优的湿润区, 据调查发现, 该区的巨柏只有几株孤立木, 鉴于该分布区与其他巨柏分布区的距离较远, 我们认为它处于孤立的特殊状态, 其分布性质有待进一步研究。
巨柏群落中共调查到维管束植物130种, 分别隶属于43科, 106属。巨柏群落植物的科、属组成分散, 与种的数目相比, 科、属数目相对较多, 尤其是属的组成最突出, 这种现象说明巨柏群落属于一个比较古老的植物区系。种子植物属的分析表明, 巨柏群落温带成分及其变型占有优势, 植物区系具有明显的北温带性质。从种的分布类型来看, 区系组成成分中含有大量的中国(西藏)特有种和中国-喜马拉雅分布种, 特有现象明显。
巨柏通常是群落的主要建群种, 形成单优群落。各巨柏种群的生长状况排序为:林芝种群 > 米林种群 > 朗县种群, 总体上由半湿润向半干旱分布区逐渐减弱。群落中灌木层和草本层植物多以全缘的草质小型单叶地面芽植物为主, 具有落叶性质。巨柏群落的结构在半湿润区分层比较明显, 至半干旱区, 随着各层的植物种类和数量减少, 林下植被逐渐稀疏, 结构趋于简单。
种群结构分析表明, 林芝种群的径级基本连续, 结构呈纺锤形, 该种群正处于成熟向老龄发展的阶段, 由于更新受阻导致种群结构失调, 其老龄化过程将会加剧; 米林种群径级分布不连续, 在Ⅱ、Ⅲ级出现断代, 更新过程在幼龄阶段被阻断, 加之上层成年个体数量的锐减, 种群扩展困难, 已呈现萎缩的态势; 朗县种群径级结构较完整, 种群的连续性和更新状况都优于林芝和朗县种群, 目前仍处于稳定状态。3个巨柏种群都有1个共同之处:在幼龄时期种群都出现了不同程度的缢缩, 更新不良造成种群比例失调, 进而导致种群扩展困难。由此推断, 巨柏的濒危原因与其更新过程受阻密切相关, 其作用机理和过程还有待进一步分析和研究。
巨柏作为一个古老的物种延续至今, 见证了青藏高原地质历史和气候的变迁, 具有极高的科学研究价值, 它能为古地理学、古植物学、古气候学等学科的研究提供极其有价值的参考。目前, 巨柏濒危的机理和过程仍不是很清楚, 对其保护力度相当不够, 多数的野外种群仍处于自生自灭状态, 未得到有效保护。鉴于目前状况, 为了有效地保护这个珍稀濒危树种, 应加强其遗传学和保护生物学的研究, 并以此为基础, 采取就地保护和迁地保护相结合的措施, 综合实施保护对策。如从不同的种群分布地收集繁殖材料, 利用现代生物技术手段培育具有遗传差别的个体, 增加巨柏的遗传变异水平和基因交流, 增加种群的异质性, 促进这一物种的进化潜能(Schemske et al., 1994; Colling et al., 2002); 进行一定规模的人工苗繁育, 在巨柏分布区和适宜区营造人工林, 促进种群更新, 增加种群的幼龄个体数量, 人为扩大种群空间。
陈端. 1994. 巨柏的育苗造林技术. 林业科技, 19(2): 6-7. |
陈端. 1995. 西藏巨柏的研究现状与前景. 西藏科技, 68(2): 7-11. |
陈端. 1996. 西藏巨柏高径生长的回归分析. 西藏科技, (2): 29-311. |
蔡飞, 宋永昌. 1997. 武夷山木荷种群结构和动态的研究. 植物生态学报, 21(2): 138-148. DOI:10.3321/j.issn:1005-264X.1997.02.005 |
于永福. 1999. 国家重点保护植物名录. 植物杂志, (5): 4-11. |
胡匡祜, 苏万芳, 李子孝, 等. 1993. 西藏巨柏的核型图像自动分析与识别的研究. 生物物理学报, 9(2): 328-333. |
黄健, 康新. 1993. 现代生境与巨柏群落的重建. 西藏科技, (1): 101. |
李文华. 1985. 西藏森林. 北京: 科学出版社, 30-149.
|
李乾振, 李朝晖. 2003. 闽东引种巨柏育苗试验初报. 林业科学, 39(1): 184-186. |
梁士楚. 1992. 贵阳喀斯特山地云贵鹅耳枥种群动态研究. 生态学报, 12(1): 53-59. DOI:10.3321/j.issn:1000-0933.1992.01.013 |
梁红平. 1990. 巨柏的核型分析. 植物学报, 32(8): 653-6561. |
曲仲湘, 吴玉树, 王焕校. 1983. 植物生态学. 北京: 高等教育出版社, 1-323.
|
武吉华, 张绅. 1995. 植物地理学. 北京: 高等教育出版社, 98-104.
|
王景升, 李衡. 2005. 西藏巨柏人工苗木生长研究. 西藏科技, (3): 47-49. DOI:10.3969/j.issn.1004-3403.2005.03.015 |
汪松, 解炎. 2004. 中国物种红色名录. 北京: 高等教育出版社, 305.
|
吴兴, 赵南先, 段代祥, 等. 2005. 西藏特有珍稀植物巨柏的研究进展与展望. 福建林业科技, 32(2): 160-164. |
吴征镒. 1983. 西藏植物志:第1卷. 北京: 科学出版社.
|
吴征镒. 1991. 中国种子植物属的分布区类型. 云南植物研究, (增刊Ⅳ): 1-139. |
中国科学院青藏高原综合科学考察队. 1984. 西藏气候. 北京: 科学出版社, 142-170.
|
中国科学院青藏高原综合科学考察队. 1988. 西藏植被. 北京: 科学出版社, 58-143.
|
壮青. 1977. 高原巨柏傲苍穹. 科学实验, (11): 5-6. |
郑万钧, 傅立国, 诚静容, 等. 1975. 中国裸子植物. 植物分类学报, 13(4): 73. |
郑万钧, 傅立国. 1978. 中国植物志:第7卷. 北京: 科学出版社, 334-335.
|
Colling G, Matthies D, Reckinger C. 2002. Population structure and establishment of the threatened long-lived perennial Scorzonera humillis inrelation toenvironment. Journal of Applied Ecology, 39: 310-320. DOI:10.1046/j.1365-2664.2002.00714.x |
Daubenmire. 1947. Plat communities-a texbook of plant synecology. Beijing: Peoples Education Press, 325-326.
|
Knowles P, Grant MC. 1983. Age and size structure analysis of engelmannspruce, ponderosapine, loadgepole pine, andlimber pine in cororado. Ecology, 64: 1-9. DOI:10.2307/1937322 |
Parker AJ, Peet R K. 1984. Sizeandage structure of coniferforests. Ecology, 65: 1685-1689. DOI:10.2307/1939148 |
Raunkiaer C. 1934. The Life forms of plants and statistical plant geography. Beijing: Oxford Clarendon Press, 632.
|
Schemske D W, Husband B C, Ruckelshaus MH, et al. 1994. Evaluating approaches tothe conservation of rare and endangered plants. Ecology, 75: 584-606. DOI:10.2307/1941718 |