 2007, Vol. 43
2007, Vol. 43文章信息
- 喻晓丽, 邸雪颖, 宋丽萍.
- Yu Xiaoli, Di Xueying, Song Liping.
- 水分胁迫对火炬树幼苗生长和生理特性的影响
- Effects of Water Stress on the Growth and Eco-physiology of Seedlings of the Rhus typhina
- 林业科学, 2007, 43(11): 57-61.
- Scientia Silvae Sinicae, 2007, 43(11): 57-61.
-
文章历史
- 收稿日期:2007-03-29
-
作者相关文章



水分胁迫诱导植物体内发生各种生理生化反应,使植物细胞生理脱水,导致植株生长停止,光合作用受抑,呼吸紊乱,整个代谢异常,导致植物受害(吴志华等,2004)。因此,近年来,对植物水分关系的探讨已成为植物生理生态学研究领域的重要课题之一(Li et al., 2000)。火炬树(Rhus typhina)是漆树科盐肤木属树种,原产北美洲。火炬树秋季花叶赤红,树形美观,最早在1959年由北京植物园作为观赏树种引入我国,1974年后向全国各省区推广,目前以黄河流域以北各省(区)栽培较多,主要用于荒山、盐碱荒地绿化兼作风景林树种(潘志刚等, 1994)。目前对火炬树已从苗木培育、发育解剖、形态结构、组织培养、克隆和生物量特征等方面进行了较多的研究(马淑英等,1997;毛学文等,1997;段新玲等,2000;庞福生等,2001;张明如等,2004),但有关火炬树的抗旱生理方面研究较少。采用人工控制施水量的方法,开展不同水分胁迫处理对火炬树幼苗生长和生理的影响的模拟试验,分析火炬树对土壤水分胁迫的生态反应,以及探讨火炬树幼苗生长对水分影响的适应机制,其结果为火炬树的推广利用提供理论依据。
1 材料和方法 1.1 试验设计2006年5月初将1年生火炬树幼苗定植于盆中(35 cm×25 cm×35 cm),每盆1株,装入土壤5 kg(含水量35.12%,最大持水量513.90 g·kg-1,田间持水量为418.56 g·kg -1),在东北林业大学林场温室大棚内进行培养,保证土壤水分充足,使之不会成为苗木生长的限制因子。于6月中旬选取长势一致的苗木,设置4个水分供应梯度,每个处理30盆,采用称重法进行控水处理,土壤含水量分别维持在:对照(CK), 土壤水分为田间最大持水量的55%~60%;轻度胁迫(T1),维持田间最大持水量的35%~40%;中度胁迫(T2),维持最大田间持水量的25 %~30%;重度胁迫(T3),维持田间最大持水量的15%~20%。6月30日达到控水梯度,每隔1 5 d于8:00—9:00从幼苗选取完全展开叶位相同的成熟叶片测定各项生理指标,共进行连续5次测定,每个处理采集3棵植株,每棵植株测定3个重复,共9个重复。生长指标在10月22日进行测定。
1.2 光合作用测定使用Licor-6400便携式光合作用测定仪,在400 μmol·L-1CO2和1 000 μmol·m-2s-1光强下测定净光合速率(Pn, μmol·m-2s-1)、蒸腾速率(Tr, mmol·m-2s-1),并计算出水分利用效率(Ew, Pn/Tr),所有测定均在9:00—11:00 25 ℃的室温环境下完成。
1.3 光合色素含量的测定叶绿素及类胡萝卜素含量的测定,采用80%丙酮提取,比色法测定(李合生,2000)。
1.4 脯氨酸和丙二醛(MDA)含量的测定丙二醛含量的测定采用硫代巴比妥酸法,游离脯氨酸含量的测定采用酸性茚三酮法(汤章城,1999)。
1.5 生长测定在2006年10月22日,每水分处理组随机选取9株幼苗,测定其株高、基径。测定每株幼苗的主根长度及第1级侧根数,最后用烘干法测得根、茎、叶干质量,并计算总生物量、根质量比(根质量/总生物量)、茎质量比(茎质量/总生物量)和叶质量比(叶质量/总生物量)。
1.6 数据处理本试验属于单因子(水分)多水平试验,取得的数据利用SPSS11.5软件系统进行ANOVA方差分析,采用Duncan法进行多重比较,并用字母法标记,图表制作采用EXCEL软件。
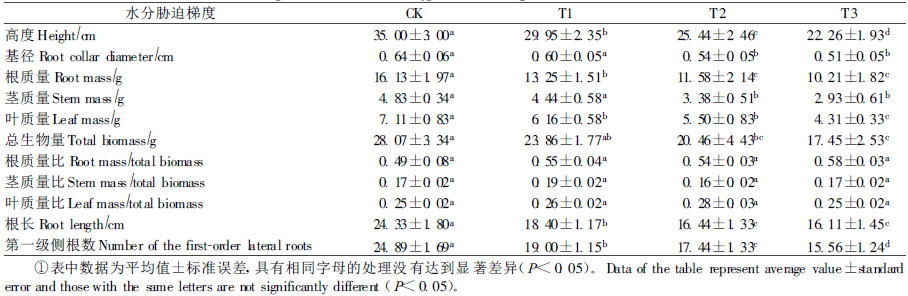
2 结果与分析 2.1 水分胁迫对生物量及分配的影响表 1表明,水分胁迫处理对火炬树幼苗高度、基径、生物量(根、径、叶以及总生物量)、根长以及第1级侧根数均有显著影响,但对根质量比、茎质量比及叶质量比的影响不显著。水分胁迫显著降低了火炬树幼苗的高生长和基径生长。CK组的火炬树幼苗株高与基径生长表现出最佳,分别为35.00 cm和0.64 cm。水分胁迫处理明显降低了火炬树幼苗的生物量积累,对火炬树总生物量影响中,CK与T2和T3组间差异显著(P<0.05),幼苗根、茎及叶质量与总生物量对水分含量变化的响应相似,随着含水量的减少而降低。而轻度水分胁迫下火炬树幼苗的茎质量和总生物量与对照组CK无显著差异(P>0.05),但T1组的根质量和叶质量显著低于对照组CK。水分胁迫对地上部分和地下部分生物量的分配影响不显著(P>0.05)。水分胁迫处理对幼苗根长和根数量的影响明显(P<0.05),随着水分胁迫程度的加剧,火炬树根长变短,第一级侧根数越少。
|
|
如图 1所示,水分梯度能显著影响火炬树幼苗光合作用,对Pn、Tr以及E w的影响均达到极显著性(P<0.05)。随着胁迫时间的延长,火炬树幼苗的Pn和Tr变化趋势基本一致,随着胁迫程度的加剧而逐渐降低,但是Pn的降低幅度低于Tr,所以3个水分处理组的WUE显著高于对照。T3组与对照组CK相比,Pn和Tr的总平均值降低了38.26%和54.09%,Ew平均值增加30.52%,表明火炬树以提高水分利用效率来适应逐渐加剧的水分胁迫条件。但是胁迫后期,T3组Ew的增幅显著低于T1和T2组,其原因可能是胁迫后期在重度水分胁迫条件下,火炬树幼苗的生长发生衰退,适应能力有所降低。

|
图 1 不同水分处理下火炬树幼苗的净光合速率、蒸腾速率和水分利用效率 Fig. 1 Net photosynthetic rate, transpiration rate and water use efficiency of Rhus typhina seedlings under different soil water stress |
不同水分处理对火炬树幼苗叶片叶绿素a(Chla)、叶绿素b(Chlb)和叶绿素总量(Chl)影响显著,而对叶绿素a/b(Chla/Chlb)影响不明显(图 2)。3个水分胁迫组的叶绿素a(Chla)、叶绿素b(Chlb)和叶绿素总量(Chl)在胁迫前期和中期均显著高于对照组CK,并随着胁迫程度的加剧而逐渐增加,但30 d后增幅降低。胁迫60 d后,均显著低于对照,T1组降低幅度最大。

|
图 2 水分胁迫对火炬树叶片叶绿素含量的影响 Fig. 2 Content of chlorophyll of Rhus typhina seedlings under different soil water stress |
在整个胁迫期间,T1、T2和T3处理组的脯氨酸含量随着胁迫程度的加剧而逐渐升高,显著高于对照CK,胁迫45 d时,增加的幅度最大,平均增幅为68.9%。胁迫后期,增加的幅度有所降低,75 d时,平均增幅为15.39%。
MDA含量随着胁迫程度的加剧而显著增加,平均增幅为39.07%。随着胁迫的进展,3个处理组的MDA含量逐渐降低,45 d时,均显著低于对照,T3组降低幅度最大。胁迫后期,3个处理组的MDA含量与对照相比又略有升高,但升高的幅度很小,60 d时平均增幅为6.07%,75 d时为16.7%。

|
图 3 不同水分处理对火炬树幼苗脯氨酸和丙二醛含量 Fig. 3 Contents of free proline and MDA in needles of Rhus typhina seedlings under different soil water stress |
研究结果表明,长期水分胁迫处理显著抑制了火炬树幼苗的生长与气体交换。在受到中度和重度水分胁迫时, 火炬树幼树叶、茎、根和总生物量的积累均受到抑制。火炬树幼苗不同器官的生物量对不同强度水分胁迫反应不同, 轻度水分胁迫对茎质量和总生物量影响不显著,而显著降低了叶质量和根质量,说明火炬树的叶片和根部生长对水分胁迫影响较敏感。水分胁迫下火炬树幼苗生长受到抑制,株高与地径生长减缓,生物量积累减小,主要原因是由于水分胁迫导致火炬树幼苗叶片Pn显著降低,从而抑制了幼苗的生长和生物量积累。随着水分胁迫的加剧下,显著降低了火炬树幼苗的根生长,主根长度变短,第一级侧根数减少,使幼苗生长受到影响,表现为各部分器官的生物量均有降低。但是对地上部分生物量和地下部分生物量的分配影响不显著,多数研究结果表明,干旱胁迫下, 幼苗会将更多的资源分配到根系生长,以便吸收更多的水分和营养物质,来提高竞争生长能力,许多植物对资源限制都做出了类似的反应(Esch et al., 1998;Xiao et al., 2002;Zhu et al., 2005),但火炬树与此研究结果不一致,其原因有待进一步研究。
在水分胁迫条件下, 火炬树幼苗的Pn和Tr均随着水分胁迫程度的加剧而降低,这与朱教君等(2005)、杨燕等(2005)对樟子松(Pinus sylvestris var. mongolica)以及杨燕等对云杉(Picea asperata)的研究结果类似。水分胁迫下火炬树Pn下降的原因可能是在水分胁迫条件下, 气孔导度减小, 导致胞间CO2浓度和蒸腾速率下降, 进而影响光合速率; 另外, 水作为光合作用的原料之一, 当其供应不足时, 也直接导致光合速率的降低。同时火炬树大幅度降低Tr,可理解为这是火炬树对水分胁迫的一种生理适应策略,以提高Ew在来适应水分胁迫。这表明火炬树在较低的土壤含水量条件下, 具有忍耐干旱胁迫的能力。
陈坤荣(1997)的研究结果表明,水分胁迫下增加叶绿素含量,增强了逆境中的加勒比松(Pinus caribaea)生存能力。高抗旱性甘薯叶绿素在水分胁迫下降幅度小于低抗旱性品种(张明生等,2001)。在水分胁迫下,3个处理组的火炬树叶绿素含量显著高于对照,可能与植物对环境因子的补偿和超补偿效应有关(邹春静等,2004)。胁迫后期,水分胁迫组的叶绿素含量低于对照,主要原因可能是干旱胁迫使叶绿体片层中的Chla/b-Pro复合体合成受到抑制(Alberte et al.,1977),或者与水分胁迫诱导叶绿体发生膜质过氧化而产生的破坏作用有关(Jiang et al., 1994)。张明生等(2001)研究表明,叶绿素a/b比值下降的程度也可以评定甘薯的抗旱性。火炬树叶绿素a/b比值在水分胁迫15 d时,各处理均高于对照,30 d后,3个处理均低于对照,但降低的幅度不是很大,可能是长时间的水分胁迫下,叶绿素a对活性氧的反应较叶绿素b敏感,使得叶绿素a的增加幅度低于叶绿素b,所以导致叶绿素a/b比值下降,从降低的幅度来看,火炬树对水分胁迫的抗性比较高。
水分胁迫下,丙二醛的积累会引起细胞膜过氧化作用,由于氧化膜的有序性降低,结构遭到了破坏,从而影响了细胞的物质代谢即吸收与同化作用(曹慧等,2001),其含量可以反映植物遭受逆境伤害的程度(Zou,2000)。本研究中,火炬树幼苗丙二醛(MDA)含量在胁迫初期增加,表明水分胁迫的逆境条件会对火炬树幼苗造成伤害,这与大部分的研究结果相似(Tükan et al., 2005;Xiong et al., 1996;Yan et al., 1999;Li et al., 2002)。胁迫中期由于前期的抗旱锻炼,3个水分处理组的丙二醛(MDA)含量显著降低,说明对水分胁迫已具有一定的适应性,在胁迫后期,可能由于火炬树生长处于衰退期,对水分胁迫的适应性有所降低,所以3个水分胁迫组与对照相比丙二醛(MDA)含量又有所增加,但增加的幅度不是很大,表明受到的伤害程度较小。
一般认为植物体内游离的脯氨酸含量不高,只有在胁迫条件下,才迅速增加(汤章城,1986)。Singh等(1972)提出在大麦中, 脯氨酸的积累与抗旱性成正相关。任文伟等(2000)对不同地种群羊草在胁迫下脯氨酸含量进行比较研究,认为羊草的抗旱性与脯氨的积累特性及其生境有着密切的联系。胁迫条件下脯氨酸含量的增加可能对防止细胞内水分的过分丢失、维持细胞膜的完整性起重要作用。在本研究中,火炬树幼苗中游离脯氨酸含量在整个干旱胁迫过程中均随着胁迫程度的加剧而呈现升高的趋势,说明游离脯氨酸在火炬树的抗旱机制中具有较大作用。
从本试验的研究结果来看,火炬树通过降低Pn、Tr和大幅度提高Ew来适应渐加剧的水分胁迫。叶绿素含量在胁迫前期和中期显著高于对照,提高了火炬树的抗旱能力。整个胁迫期间,游离脯氨酸在水分胁迫下呈显著升高趋势,在火炬树的抗旱机制中具有重要作用。MDA含量在胁迫前期显著升高,说明火炬树对水分胁迫较敏感,胁迫中期3个处理组的MDA含量显著降低,表明火炬树对不同水分胁迫程度具有了一定的适应能力。在衰退期其适应能力有所下降,但MDA含量的增幅不大,其伤害程度较小。
陈坤荣, 王永义. 1997. 加勒比松耐旱性生理特征研究. 西南林学院学报, 17(4): 9-15. |
曹慧, 王孝威, 曹琴, 等. 2001. 水分胁迫下新红星苹果超氧物自由基累积和膜脂过氧化作用. 果树学报, (4): 10-13. |
段新玲, 任东岁, 赵树珍. 2000. 火炬树的组织培养及植株再生. 植物生理学通讯, 36(6): 535. |
李合生. 2000. 植物生理生化试验原理和技术. 北京: 高等教育出版社, 134-200.
|
马淑英, 胡正海. 1997. 火炬树分泌道的发育解剖学研究. 西北植物学报, 17(5): 112-117. DOI:10.3321/j.issn:1000-4025.1997.05.022 |
毛学文, 施文甫, 王弋博. 1997. 火炬树腺毛的形态结构和发育的研究. 西北植物学报, 17(6): 137-139. DOI:10.3321/j.issn:1000-4025.1997.06.028 |
潘志刚, 游应天. 1994. 中国主要外来树种引种栽培. 北京: 北京科学技术出版社, 525-528.
|
庞福生, 刘孝智, 李晓伟, 等. 2001. 侧柏火炬树混交林蓄水保水效益的研究. 水土保持研究, 8(3): 14-15. DOI:10.3969/j.issn.1005-3409.2001.03.005 |
任文伟, 钱吉, 郑师章. 2000. 不同地理种群羊草在聚乙二醇胁迫下含水量和游离脯氨酸含量的比较. 生态学报, 20(2): 349-352. DOI:10.3321/j.issn:1000-0933.2000.02.028 |
汤章城. 1986. 水分胁迫和植物的气孔运动. 植物生理生化进展, (4): 43-50. |
汤章城. 1999. 现代植物生理学实验指南. 北京: 科学出版社, 302-306.
|
吴志华, 曾富华, 马生健, 等. 2004. 水分胁迫下植物活性氧代谢研究进展(综述Ⅰ). 亚热带植物科学, 33(2): 77-80. DOI:10.3969/j.issn.1009-7791.2004.02.023 |
杨燕, 刘庆, 林波, 等. 2005. 不同施水量对云杉幼苗生长和生理生态特征的影响. 生态学报, 25(9): 2152-2158. DOI:10.3321/j.issn:1000-0933.2005.09.004 |
张明生, 谈锋. 2001. 水分胁迫下甘薯叶绿素a/b比值的变化及其与抗旱性的关系. 种子, 20(4): 23-25. |
邹春静, 韩士杰, 徐文铎, 等. 2004. 沙地云杉生态型对干旱胁迫的生理生态响应. 应用生态学报, 14(9): 1446-1450. |
朱教君, 康宏樟, 李智辉, 等. 2005. 水分胁迫对不同年龄沙地樟子松幼苗存活与光合特性影响. 生态学报, 25(10): 2527-2533. DOI:10.3321/j.issn:1000-0933.2005.10.010 |
张明如, 翟明普, 贾黎明, 等. 2004. 火炬树克隆植株生长和生物量特征的研究. 林业科学, 40(3): 39-45. DOI:10.3321/j.issn:1001-7488.2004.03.006 |
Alberte R S, Thomber J P, Fiscs E L. 1977. Water stress effects on the content and organization of chlorophyll in mesophyll and bundle sheach chloroplasts of maize. Plant Physiol, 58: 351-353. |
Esch A, Mengel K. 1998. Combined effects of acid mist and frost drought on the water status of young spruce trees(Picea sbies). Envir and Expr Bot, 39(1): 57-65. DOI:10.1016/S0098-8472(97)00035-X |
Jiang M Y, Yang W Y, Xu J, et al. 1994. Active oxygen damage effect of chlorophyll degradation in rice seedlings under osmotic stress. Acta Botanica Sinica, 36(4): 289-295. |
Li H J, Chai B F, Wang M B. 2000. Study on the water physio-ecolgical characteristics of Populous Beijingensis. Acta Ecological Sinica, 20(3): 417-422. |
Li M, Wang G X. 2002. Effects of drought stress on activities of cell defense enzymes and lipid peroxidation in Glycyrrhiza uralensis seedlings. Acta Ecologica Sinica, 22(4): 503-507. |
Singh T N, Aspinall D, Palag L G. 1972. Proline accumulation and varietal adaptability to drought in barley A potential metabolic meature of drought resistance. Nature New Boil, 236: 188-190. |
Tükan I, Bor M, Ozdemir F, et al. 2005. ifferential responses of lipid peroxidation and antioxidantrs in the leaves of drought-tolerant P.acutifolius Gray and drought-sensitive P.vulgaris L.subjected to polyethylene glycol mediated water stress. Plant Science, 168: 223-231. DOI:10.1016/j.plantsci.2004.07.032 |
Xiong D J, Lin Z H, Yang B Y, et al. 1996. Studies of four kinds of isozymes and the changes of content of malond ialdehyde in maize in water-logging or chilling stress. Journal Nanchang University: Natural Science, 20(4): 314-319. |
Xiao C W, Zhou G S, Ma F Y. 2002. Effect of water supply changge on morphology and growth of dominant plants in Maowusu sandland. Acta Phytoecologica Sinica, 26(1): 69-76. |
Yan X F, Li J, Zu Y G. 1999. Effect of drought stress on acticity of cell defense enzymes and lipid peroxidation in korean pine seedlings. Acta Ecologica Sinica, 19(6): 850-854. |
Zou Q. 2000. Plant physiological experimental guidance. Beijing: Chinese Agricultural Press.
|
Zhu X W, Huang Z Y, Zhang S M, et al. 2005. The responses of seed germination, seedling emergence and seedling growth in Agropyron cristatum to sand water content in Otindag Sandland, China. Acta Ecologica Sinica, 25(2): 364-370. |