 2007, Vol. 43
2007, Vol. 43文章信息
- 王秀伟, 毛子军.
- Wang Xiuwei, Mao Zijun.
- 兴安落叶松人工林冠层气体交换的时空特性
- Temporal and Spatial Variation in Gas Exchange in Canopy of Larix gmelinii Plantation
- 林业科学, 2007, 43(11): 43-49.
- Scientia Silvae Sinicae, 2007, 43(11): 43-49.
-
文章历史
- 收稿日期:2007-08-06
-
作者相关文章



森林生态系统的碳平衡主要是由2个通量决定的,即光合作用的碳获得和呼吸作用的碳释放(Turnbull et al., 2003)。这2种通量决定于树种本身的生物学特性和环境条件。研究表明,光合作用与呼吸作用对环境变化的反应是独立的,因此非线性地影响森林的碳获得(Ryan, 1991; Dewar et al., 1999; Gunderson et al., 2000)。在树木个体水平上,冠层的光合能力和呼吸活性的分布是变化的(Field, 1983; Evans, 1989; Anten et al., 1995; Hollinger, 1996)。通常冠层顶部比底部叶片的光合能力强20%~40%(Meir et al., 2002; Niinemets et al., 2006)。对冠层水平上呼吸作用的研究发现,树冠上层比下层的叶片呼吸作用强,可能是由于上层叶片的光合器官具有较强活性而需要的维持呼吸较高(Griffin et al., 2002; Whitehead et al., 2004)。然而,在自然条件下,植物叶片暴露于许多环境因子中,这些环境因子可能影响呼吸速率对温度的反应方式(Turnbull et al., 2001;2003)。所以,在不同冠层中,光合作用和呼吸作用等生理过程对温度和光照反应的研究还很不全面(Xu et al., 2006)。
兴安落叶松(Larix gmelinii)人工林占东北地区人工林面积的70%,仅黑龙江省就有落叶松人工林84.08万hm2,占全省人工林总面积的80%以上。我国对兴安落叶松林生产力及碳平衡的研究主要集中在天然林(王玉辉等,2001;蒋延玲等,2002), 而对人工林生产力和碳平衡的研究甚少,尤其是在生理水平上研究落叶松人工林冠层的光合生产及碳平衡还非常有限。
为了更准确地估测落叶松人工林生态系统的光合生产力和碳平衡,对树木个体光合作用和呼吸作用在时间和空间上的变化式样及其与温度和光照的关系进行详细研究是非常必要的。本研究在于揭示兴安落叶松人工林冠层水平上光合作用和呼吸作用在时间和空间上的变化式样及其与温度和光照的关系,为准确估测兴安落叶松人工林碳平衡提供理论依据。
1 材料与方法 1.1 样地设置研究对象为34年生兴安落叶松人工林,由于混有少量的白桦(Betula platyphylla)和水曲柳(Fraxinus mandshurica),所以冠层主要由兴安落叶松、白桦和水曲柳3树种组成。该林分位于东北林业大学帽儿山林场老山生态定位站,地处长白山系张广才岭西北部小岭余脉,地理坐标为127°34′ E,45°20′ N,平均海拔340 m。该地区地带性土壤为典型暗棕壤,年平均气温2.8 ℃,年平均相对湿度70%,年平均降水量723.8 mm,年平均蒸发量1 093.9 mm,年平均日照数2 471.3 h,无霜期120~140 d。兴安落叶松和白桦于5月中旬展叶,10月初开始落叶。
2004年7月在兴安落叶松人工林内设立了40 m×50 m的标准地,并进行了样地调查,兴安落叶松平均树高22.2 m±4.29 m,平均胸径20.3 cm±5.73 cm;白桦平均树高21.0± 8.64 m,平均胸径19.1 cm±8.71 cm。
1.2 试验方法 1.2.1 光合速率的测定野外测定于2005年生长季6、7、8、9月的月初进行,在样地内设有一个22 m高的观测塔,光合速率和呼吸速率均在塔上进行活体测定。在观测塔周围,根据林分平均胸径、平均树高、平均冠幅和冠长, 选取白桦和兴安落叶松各1株标准木,将树冠等分为上、中、下3层(与地面距离下层14~16 m、中层16~18 m、上层18~21 m),白桦每层取3片有代表性的叶片;对兴安落叶松,则在每层侧枝上选择代表性的当年生枝、往年生枝上各3束针叶, 因为针叶在当年生小枝上轮生,而且枝内也含有叶绿素,同样进行光合作用,所以测定当年生枝针叶光合速率时是连同其着生的嫩枝一同测定的,而在往年生枝针叶光合速率测量时只测针叶束。
采用便携式光合作用系统(Li-6400,LI-COR, Nebraska, USA)在晴朗天气的上午选择树冠东南方向、生长状态一致的健康叶片,测定2 000 μmol·m-2s-1光强下的净光合速率(Pn)及0光强下的暗呼吸速率,每次测定取值前稳定15~20 min左右。由于兴安落叶松的叶片为针叶,不能充满叶室,测量光合速率后将针叶取回,用叶面积仪(Li-3000,LI-COR, Nebraska, USA)测定其叶面积,重复测量3次取平均值,然后采用软件(Li-6400 fluorescence and gas exchange system)对测量值进行重新计算。测定在自然条件下进行,白桦的叶片温度为16~29 ℃, 叶室内湿度59%~73%。落叶松叶片温度为16~30 ℃;叶室内湿度为61%~73%。
1.2.2 呼吸速率与温度的拟合呼吸速率对温度的响应采用Arrhenius曲线拟合(Lloyd et al., 1994):
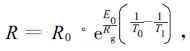
|
(1) |
式中:R0为基础温度下的呼吸速率(落叶松当年生枝针叶14.67 ℃,落叶松往年生枝针叶14.56 ℃, 白桦13.83 ℃),Rg是气体常数(8.314 J·mol-1K-1);Eo是与活化能相关的一个参数,表征对温度反应的程度。
1.2.3 叶面积指数叶面积指数使用叶面积指数仪(LAI-2000, LI-COR, Nebraska, USA)每半个月测定1次。
1.2.4 消光系数的计算如果把叶层看作是均匀的,则光强度减弱可以按M. Monsi和T. Saeki修正的消光方程式(王伯荪,1987),即
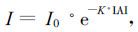
|
(2) |
式中:I为距植物冠层顶端一定距离的光强度;I0树冠顶端的入射光强度;K为该特定群落的消光系数(light extinction coefficient);LAI为叶面积指数。
1.2.5 数据处理方差分析、回归分析及曲线回归采用统计软件SPSS 11.5 (SPSS Inc, USA)。
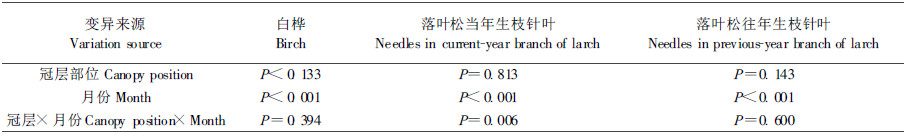
2 结果与分析 2.1 光合作用白桦最大净光合速率在冠层和月份间的差异均显著,其交互作用不显著,因此对白桦最大净光合速率冠层和月份间的差异分别进行了分析;落叶松当年生枝针叶最大净光合速率在冠层和月份间及其交互作用的差异均显著(表 1),因此对同一冠层不同月份和同月份不同冠层落叶松当年生枝针叶最大净光合速率进行了比较分析;落叶松往年生枝针叶最大净光合速率在冠层和月份间及其交互作用的差异均不显著。
|
|

白桦最大净光合速率在冠层上、中、下3层之间差异均显著,上层的最大净光合速率最高,达(17.56±0.62) μmolCO2·m-2s-1。在整个生长季中,上层较中层平均高出34.95%,比下层高83.36%,中层比下层高出35.87%(图 1,A);净光合速率的最大值出现在7月,为(15.55±0.71) μmolCO2·m-2s-1。在整个生长季中,最大和最小的月最大净光合速率平均值相差4.39 μmolCO2·m-2s-1 (图 1,B)。

|
图 1 白桦、落叶松不同冠层、月份最大净光合速率 Fig. 1 Pnmax of birch and larch through the canopy and during the growth season A.白桦不同冠层位置上最大净光合速率在整个生长季(6—9月份)的平均值;B.白桦整个冠层(冠层上、中、下3层最大净光合速率的平均值)生长季内的最大净光合速率(a、b、c表示P=0.05显著性差异组别);C.落叶松当年生枝针叶不同月份最大净光合速率(a1、b1表示相同月份不同冠层位置间P=0.05显著性差异组别;a2、b2表示相同冠层位置不同月份间P=0.05显著性差异组别);D.白桦、兴安落叶松当年生枝针叶和往年生枝针叶生长季中最大净光合速率动态。图中为均值(±SEM),t检验n=12(A),9(B)和3(C)。不同字母表示0.05水平上差异显著。 A. Mean values of Pnmax in different canopy positions of birch during the growth season (Jun.—Sep.); B. Dynamics of Pnmax in canopy level of birch (means of upper, middle and lower layers) during the growth season(a, b, c, significant difference at P=0.05); C. Pnmax of larch in differe nt canopy positions during the growth season (a1, b1, different canopy position in the same month at P=0.05; a2, b2, dif ferent month in the same canopy position at P=0.05); D. Dynamics of Pnmax of birch leaves, current-year and previous-year branch needles of larch. Values shown are means (±SEM) where n=12 (A), n=9 (B), n=3(C). Mean s were compared in pairs by t-test. If two values are followed by the same letter, they are not significantly different at P=0.05. |
落叶松往年生枝针叶最大净光合速率在冠层和月份间的差异均不显著,其平均光合速率为8.22 μmolCO2·m-2s-1。
落叶松当年生枝针叶6、7月份最大净光合速率在冠层的上、中、下3层之间的差别不显著(图 1C);8月份树冠上层的最大净光合速率与中层和下层差异显著,上层的当年生枝针叶最大净光合速率最高,为(13.09±0.97) μmolCO2·m-2s-1,比中层高51.05%,比下层高44.32%,中层和下层的差异不显著;9月树冠的最大净光合速率从上到下逐渐降低,上层为(13.64±3.02) μmolCO2·m-2s-1,平均比中部高35.18%,比下部高93.74%。在同一冠层上,落叶松当年生枝针叶在不同月份的最大净光合速率也存在着差异,上层当年生枝针叶最大净光合速率在8月和9月要比6月和7月高102.25%;中层6、7、8、9月间的差异不显著;下层6月份的最大净光合速率最低,与8月、9月差异显著,8月比6月高66.25%,9月比6月高29.05%,而7月与6、8、9月的差异没有达到显著的水平(图 1C)。
2.2 呼吸作用对白桦和落叶松针叶的暗呼吸速率月份间和种间(白桦叶片、落叶松当年生枝和往年生枝针叶)的差异进行了比较,白桦叶片和落叶松针叶的呼吸速率在冠层间的变化均不显著,而月份间的差异显著(表 2)。
|
|
白桦的暗呼吸速率在整个生长季中呈先下降(7月)、后升高(8月)、再下降(9月)的趋势。8月份值最高,9月最低为(0.48±0.03) μmolCO2·m-2s-1(图 2)。

|
图 2 白桦叶片和兴安落叶松当年生枝针叶和往年生枝针叶暗呼吸速率季节动态 Fig. 2 Dynamics of dark respiration (Rd) of Birch leaves, current-year branch needles and previous-year branch needles of Larch during the growth season |

|
图 3 帽儿山34年生兴安落叶松人工林生长季消光系数 Fig. 3 Light extinction coefficients in 34-year-old larch plantation in Maoershan mountain |
落叶松当年生枝和往年生枝在整个生长季中的变化式样相似。落叶松当年生枝针叶暗呼吸速率在6、7、8月间的差异不显著,分别比9月(0.83±0.10) μmolCO2·m-2s-1高75.90%、78.31%和104.82%(图 2)。
落叶松往年生枝针叶8月暗呼吸速率最高为(1.63±0.10) μmolCO2·m-2s- 1,比6、7、9月分别高92.16%、83.98%、138.64%,暗呼吸速率的变化与测量时的叶片温度变化趋势一致(图 2)。
2.3 温度和光照对光合速率和暗呼吸速率的影响冠层结构的异质性引起冠层内环境的空间变化, 而叶片对此环境的适应结果会使光合构件及其相应的生理生态过程发生变化;时间的变化也同样引起环境的变化,而且叶片的不同发育阶段其光合速率和呼吸速率也发生变化;而影响光合速率和呼吸速率的主要环境因子是温度和光合有效辐射。
2.3.1 光合有效辐射和温度对光合速率的影响光合有效辐射和温度对光合速率的影响冠层位置和月份的变化对白桦和落叶松最大光合速率均有不同程度的影响。为了研究光合作用与光合有效辐射的关系, 通过测定冠层的叶面积指数, 计算其消光系数。结果表明, 试验林分冠层的消光系数在6 、7 、8 、9 月份分别为0.71 ±0.01, 0.86±0.01, 0.93 ±0.02, 0.91 ±0.02(图 3)。即生长季初期冠层的消光系数较小, 随着叶片的发育, 到8 月份达到最大, 9 月稍有降低。结合白桦、落叶松不同冠层和不同月份的最大净光合速率, 可见光照对这2 个种光合作用的影响不同。冠层位置在生长季的所有月份对白桦叶片的最大净光合速率的影响均十分显著(图 1, A), 随着冠层高度的下降光合能力逐渐降低, 说明冠层不同高度叶片光合能力的变化与冠层从上到下光量子密度(PPFD)逐渐降低有关(Ellsworth et al., 1993;Kull et al., 1998)。
落叶松当年生枝针叶的最大净光合速率在6月和7 月初不同冠层之间没有显著差别, 而在8月和9 月初上层叶片最大净光合速率要显著高于中层和下层(图 1C), 明显呈现出与光合有效辐射的正相关。而落叶松往年生枝针叶的最大净光合速率在冠层和月份之间无显著变化。
叶片最大净光合速率与叶片温度之间的相关性分析表明,白桦冠层只有上层叶片最大净光合速率与叶片温度的相关性较大(R2=0.42)(图 4,A), 其他的相关性都较差。落叶松的当年生枝(图 4,B)和往年生枝(图 4,C)的各冠层最大净光合速率与温度的相关性均很小,说明温度对两树种最大净光合速率的影响不显著。

|
图 4 叶片温度与最大净光合速率(A, B, C)、呼吸速率(D)的关系 Fig. 4 Relationship between temperature and maximum net photosynthetic rate (Pnmax) and dark respiration (Rd) during the growth season for leaves |
如前所述,白桦叶片与落叶松针叶呼吸速率在冠层间的差异均不显著,而月份间的差异显著,其主要原因之一可能是由于季节变化引起气温发生改变。所以对整个生长季的呼吸速率与叶片温度采用Arrhenius方程进行拟合分析(图 4D)。
从图 4D可见,白桦叶片和落叶松往年生枝针叶的暗呼吸速率与叶片温度拟合的决定系数均很高,分别为0.57、0.69,说明温度是影响白桦叶片和落叶松往年生枝针叶的暗呼吸速率的主要因素。落叶松当年生枝针叶的暗呼吸速率与叶片温度拟合的决定系数则低于前二者,为0.34。表明落叶松当年生枝针叶的暗呼吸速率的季节变化并不完全由叶片温度决定,可能还与叶片和幼枝的发育阶段及其他环境因素有关。
3 讨论从叶片水平来看,与落叶松相比,白桦最大净光合速率较高,在生长旺盛的6、7月份,其最大净光合速率平均约为落叶松的2倍(图 1D)。这一结果与Brodribb等(2005)对蕨类植物、裸子植物和被子植物叶片的导水率与最大净光合速率之间的关系研究结果一致,即在上述三大功能群植物中,具有导管的被子植物的光合能力比具有管胞的裸子植物要高。白桦为阔叶先锋树种,是典型的被子植物,具有喜光和生长迅速的特点,这也与其较高的光合能力有关。
在冠层的水平上,一些研究表明,植物叶片最大净光合速率从上层到下层依次递减(张小全等,2000;霍宏等,2007),分析可能与光照强度在树冠内部的垂直梯度变化直接相关。Lloyd等(2002)对西伯利亚的欧洲赤松(Pinus sylvestris)林研究表明,不同冠层光合能力的变化主要依赖于光合有效辐射。树冠不同部位由于接受的光强不同而具有不同的潜在光合能力(Stenberg, 1998)。在本研究中,白桦冠层的最大净光合速率的变化就呈现这种明显的梯度变化(图 1A),反映了该种较强的光依赖关系。但是对兴安落叶松而言,只有当年生枝针叶在生长季的后期(8、9月),冠层梯度上才有差别(图 1C),而往年生枝针叶最大净光合速率在冠层间的变化不明显。这可能与该种本身的生理、生物学特性有关,即当年生枝上的针叶的光合器官发育特点、结构及其生理过程可能与往年生枝针叶有所不同,同时可能也与具有光合能力的当年生幼枝的光合生理活动有关,因为针叶在当年生小枝上轮生,所以光合速率测定是同其着生的嫩枝一同测定的。上述这些可能的因素,需要进一步深入研究。另外,落叶松当年生枝与往年生枝针叶最大净光合速率的月动态变化有所差别,往年生针叶的Pnmax除在生长季初期稍低之外,其他月份的变化不大。而当年生枝针叶的Pnmax则在中后期(8,9月份)显著升高(图 1D),几乎达到6、7月份的2倍。这与当年生针叶的发育阶段有关,因为落叶松当年枝在整个生长季都在不断生长,光合能力也不断加强。而往年生枝针叶自展叶后很短时间就趋于稳定,不再生长。
一些研究表明,树冠不同高度叶片的呼吸速率可能不同。Xu等(2006)报道,红槲栎(Qu ercus rubra)林冠上层叶片的呼吸速率高于林冠下层,推测与光照条件、水分状况、生理过程有关。本研究表明,冠层高度对白桦叶片和落叶松枝针叶暗呼吸速率影响并不显著,而月份间的差异显著,说明光照强度在树冠内部的垂直梯度不是影响暗呼吸速率的决定因素,而温度则是其主要因子,因为树木的呼吸作用变化式样与相应生长季叶温的变化式样非常吻合(图 2)。但是温度对白桦叶片和落叶松往年生枝针叶的影响较对落叶松当年生枝针叶的影响显著,其原因可能是落叶松当年生枝针处于生长旺盛时期,在不断发育过程中,不同发育阶段其呼吸速率不同,而不是完全取决于温度。可见叶片呼吸作用的大小取决于各种复杂因素的综合作用。而光合作用与呼吸作用对环境变化的反应是独立的,这与Ryan(1991)对16种植物和Gunderson等(2000)对糖槭(Acer saccharum)的研究结果以及Dewar等(1999)采用模型估计的结果一致。
霍宏, 王传宽. 2007. 冠层部位和叶龄对红松光合蒸腾特性的影响. 应用生态学报, 18(6): 1181-1186. DOI:10.3321/j.issn:1001-9332.2007.06.002 |
蒋延玲, 周广胜. 2002. 兴安落叶松林碳平衡及管理活动影响研究. 植物生态学报, 26(3): 317-322. DOI:10.3321/j.issn:1005-264X.2002.03.010 |
王玉辉, 周广胜, 蒋延玲. 2001. 兴安落叶松林生产力模拟及其生态效益评估. 应用生态学报, 12(5): 648-652. DOI:10.3321/j.issn:1001-9332.2001.05.002 |
王伯荪. 1987. 植物群落学. 北京: 高等教育出版社, 93-94.
|
张小全, 徐德应. 2001. 18年生杉木不同部位和叶龄针叶光响应研究. 生态学报, 21(3): 409-414. |
Anten N P R, Schieving F, Werger M J A. 1995. Patterns of light and nitrogen distribution in relation to whole canopy carbon gain in C3 and C4 mono- and dicotyledonous species. Oecologia, 101: 504-513. DOI:10.1007/BF00329431 |
Brodribb T J, Holbrook N M, Zwieniecki M A, et al. 2005. Leaf hydraulic capacity in ferns, conifers and angiosperms: impacts on photosynthetic maxima. New Phytologist, 165: 839-846. |
Dewar R C, Medlyn B E, McMurtrie R E. 1999. Acclimation of the respiration photosynthesis ratio to temperature: insights from a model. Global Change Biology, 5: 615-622. DOI:10.1046/j.1365-2486.1999.00253.x |
Ellsworth S, Reich P. 1993. Canopy structure and vertical patterns of photosynthesis and related leaf traits in a deciduous forest. Oecologia, 96: 169-178. DOI:10.1007/BF00317729 |
Evans J R. 1989. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia, 78: 9-19. DOI:10.1007/BF00377192 |
Field C. 1983. Allocating leaf nitrogen for the maximization of carbon gain: leaf age as a control on the allocation program. Oecologia, 56: 341-347. DOI:10.1007/BF00379710 |
Griffin K L, Tissue D T, Turnbull M H, et al. 2001. Leaf dark respiration as a function of canopy position in Nothofagus fusca trees grown at ambient and elevated CO2 partial pressures for 5 years. Functional Ecology, 15: 497-505. DOI:10.1046/j.0269-8463.2001.00539.x |
Griffin K L, Turnbull M, Murthy R. 2002. Canopy position affects the temperature response of leaf respiration in Populus deltoides. New Phytologist, 154: 609-619. DOI:10.1046/j.1469-8137.2002.00410.x |
Gunderson C A, Norby R J, Wullschleger S D. 2000. Acclimation of photosynthesis and respiration to simulated climatic warming in northern and southern populations of Acer saccharum: laboratory and field evidence. Tree Physiology, 20: 87-96. DOI:10.1093/treephys/20.2.87 |
Hollinger D Y. 1996. Optimality and nitrogen allocation in a tree canopy. Tree Physiology, 16: 627-634. DOI:10.1093/treephys/16.7.627 |
Kull O, Niinemets Ü. 1998. Distribution of leaf photosynthetic properties in tree canopies: comparison of species with different shade tolerance. Functional Ecology, 12: 472-479. DOI:10.1046/j.1365-2435.1998.00214.x |
Lloyd J, Taylor J A. 1994. On the temperature-dependence of soil respiration. Functional Ecology, 8: 315-323. DOI:10.2307/2389824 |
Lloyd J, Shibistova O, Zolotoukhine D, et al. 2002. Seasonal and annual variations in the photosynthetic productivity and carbon balance of a central Siberian pine forest. Tellus, 54B: 590-610. |
Meir P, Kruijt B, Broadmeadow M, et al. 2002. Acclimation of photosynthetic capacity to irradiance in tree canopies in relation to leaf nitrogen concentration and leaf mass per unit area. Plant, Cell and Environment, 25: 343-357. DOI:10.1046/j.0016-8025.2001.00811.x |
Niinemets Ü, Cescatti A, Rodeghiero M, et al. 2006. Complex adjustments of photosynthetic capacity and internal mesophyll conductance to current and previous light availabilities and leaf age in Mediterranean evergreen species Quercus ilex . Plant, Cell and Environment, 29: 1159-1178. DOI:10.1111/j.1365-3040.2006.01499.x |
Ryan M G. 1991. Effects of climate change on plant respiration. Ecological Applications, 1: 157-167. DOI:10.2307/1941808 |
Stenberg P. 1998. Implications of shoot structure on the rate of photosynthesis at different levels in a coniferous canopy using a model incorporating grouping and penumbra. Functional Ecology, 12: 82-91. DOI:10.1046/j.1365-2435.1998.00169.x |
Turnbull M H, Whitehead D, Tissue D T, et al. 2001. Responses of leaf respiration to temperature and leaf characteristics in three deciduous tree species vary with site water availability. Tree Physiology, 21: 571-578. DOI:10.1093/treephys/21.9.571 |
Turnbull M H, Whitehead D, Tissue D T, et al. 2003. Scaling foliar respiration in two contrasting forest canopies. Functional Ecology, 17: 101-114. DOI:10.1046/j.1365-2435.2003.00713.x |
Whitehead D, Griffin K L, Turnbull M H, et al. 2004. Response of total night-time respiration to differences in total daily photosynthesis for leaves in a Quercus rubra L. canopy: implications for modelling canopy CO2 exchange. Global Change Biology, 10: 925-938. |
Xu C Y, Griffin K L. 2006. Seasonal variation in the temperature response of leaf respiration in Quercus rubra: foliage respiration and leaf properties. Functional Ecology, 20: 778-789. DOI:10.1111/j.1365-2435.2006.01161.x |