 2007, Vol. 43
2007, Vol. 43文章信息
- 江泽慧, 范少辉, 冯慧想, 张群, 刘广路, 宗亦尘.
- Jiang Zehui, Fan Shaohui, Feng Huixiang, Zhang Qun, Liu Guang lu, Zong Yichen.
- 华北沙地小黑杨人工林生物量及其分配规律
- Biomass and Distribution Patterns of Populus × xiaohei Plantation in Sandy Land of North China
- 林业科学, 2007, 43(11): 15-20.
- Scientia Silvae Sinicae, 2007, 43(11): 15-20.
-
文章历史
- 收稿日期:2006-10-08
-
作者相关文章



2. 中国林业科学研究院林业研究 北京 100091;
3. 国际竹藤网络中心国家林业局竹藤科学与技术重点实验室 北京 100102
2. Research Inst itute of Forestry, CAF Beijing 100091;
3. International Centre of Bamboo and Rattan Key Laboratory of Bamboo and Rattan, State Forestry Administration Beijing 100102
小黑杨(Populus × xiaohei)具有生长迅速、抗寒性强、抗旱效果好、抗病虫、适应性较广、树干通直圆满、材质洁白等优良特性(赵天锡等,1994),现已成为营造杨树速生丰产人工林的优良树种之一。目前, 国内外对杨树的研究涉及到杨树生理生态特性对气候变化的响应、转基因技术及经营措施、密度、基因型、树冠结构、CO2浓度、立地条件等因子对杨树生物量的影响等方面(Jorg et al., 2003;Victoria et al., 2005;Long et al., 2004;Anna et al., 2000;朱春全等,1997;崔浪军等,2003;Norby et al., 1999),有关小黑杨林分生物量的研究很少。对小黑杨人工林的生物量进行研究,可以揭示小黑杨生态系统能量平衡、能量流动和养分循环等功能过程的变化规律,为生态系统的碳汇和碳素循环研究提供关键数据(Houghtou et al., 2001;Liu et al., 1999;Baldwin et al., 2001;Gower et al., 1997;Kurz et al., 1999),有利于实现利用现代遥感手段预测、监测和经营森林,为木材性质评价提供基础数据。
1 试验地概况试验地设在山西省朔州市杨树丰产林实验局薛家庄国营林场(112°06′E,39°02′N)。半干旱草原气候,冬季漫长而寒冷干燥,夏季短暂而温热多雨,春秋凉爽。气温日差和年差均大,大风多,日照充足。年均温7.5 ℃左右,无霜期134 d,降水集中在6—8月,年降水量360~460 mm。主要气象灾害是干旱。平均海拔1 060~1 400 m,主要地貌为冲积平原,成土母质以马兰黄土和冲积物为主,其次为风积物,还有少量的玄武岩。土壤类型为草甸土,多呈碱性,其pH值为8~9。
调查对象为小黑杨人工林,1977年栽植,造林面积21.6 hm2,全面机耕,采用“三大一深”栽植方法,造林用苗均为二根、二杆一级苗,植穴1 m×1 m×1 m,常规经营。
2 研究方法 2.1 样地选择与调查2004年6月,根据各种密度的生长状况,在试验区内选取了3种密度(1 000、500和250株·h m-2)进行标准地设置,标准地大小为25 m × 25 m,做3个重复,共设9块标准地。每木检尺,测树高、枝下高和冠幅,并进行林下植被、土壤剖面及生物量调查。在每木检尺的基础上,以2 cm为1个径阶选取14~28 cm共8个径阶的标准木,选取株数分别为1、2、3、2、3、3、3、1,共18株做生物量调查。2006年7月对以上标准地林木进行了复查。
2.2 林木各器官生物量的测定伐倒标准木,分别秤量树干、树皮、树枝和树叶的鲜质量,然后各取一部分样品带回实验室,105 ℃烘干至恒重,计算出各器官的含水率,推算出标准木地上部分各器官的干质量;将其根系全部挖出,秤其鲜质量,取一部分样品带回实验室105 ℃烘干至恒重,推算出解析木地下部分的生物量。
2.3 生物量模型的建立本研究利用标准木胸径和各器官生物量的数据,建立小黑杨各器官生物量(W)与胸径(D)的指数回归模型W=aDb来估计林分的生物量。由于按林木各分量(干、枝、皮、叶、根)分别建立模型,模型确定后根据各分量实际观测数据分别拟合各自方程中的参数,即各分量之间干质量的估计都是独立进行的。因而造成各维量模型间不相容,即树干、树皮、树枝、树叶、树根等部位的干质量之和不等于总量。应用线性联立方程组的方法可以很好地解决模型的相容性问题(张会儒等,1999;唐守正等,2000),本研究采用线性联立方程组1:1级比例平差法来解决模型的相容性问题(唐守正等,2000)。
2.4 林分生物量的测定将标准地各株林木的胸径值代入生物量指数回归模型中,计算出林分生物量。
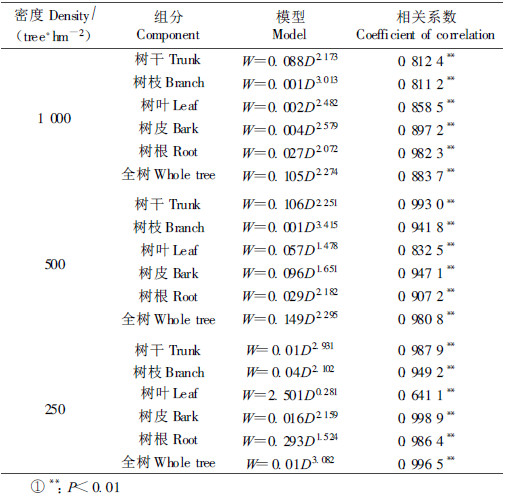
3 结果与分析 3.1 生物量模型及其相关系数由表 1可知,除250株·hm-2的小黑杨叶的生物量模型相关系数较小外(0.641 1),其余的相关系数均很高,经F检验,模型均达到极显著水平,表明模型具有良好的相关性,均可用于计算小黑杨人工林的生物量。
|
|
通过对27年生和29年生小黑杨人工林生物量的研究可知,2年间生物量增长幅度很小(表 2),按密度高低分别增长了3.32、2.30和4.20 t·hm-2,反映了27年生小黑杨人工林已经进入成熟期,生长速度缓慢。密度对器官间生物量分配有重要影响,在密度为1 000和250株·hm-2的林分中,器官间的生物量排列顺序是干>根>枝>皮>叶;在500株· hm-2的林分中,器官的生物量排列顺序是干>枝>根>皮>叶,27年生时分别为57.79、21.44、12.72、7.92和2.73 t·hm-2,29年生时分别为58.98、22.14、12.97、8.04和2.77 t·hm-2。器官生物量所占比例在同一密度不同林龄的林分中差异很小,但在不同密度的林分中差异较大。干生物量所占比例在3种密度林分中的排列顺序是1 000>250>500株·hm-2的林分,27年生时分别为63.34%、59.95%和56.32%,29年生时分别为63.28%、60.47%和56.22%;枝生物量所占比例的排列顺序是500>250>1 000株·hm-2的林分,27年生时分别为20.90%、14.92%和8.94%,29年生时分别为21.1 1%、14.81%和9.00%;叶和皮生物量所占比例的排列顺序是1 000>500>250株·hm- 2的林分,27年生时叶生物量所占比例分别为3.63%、2.66%和2.11%,皮生物量所占比例分别为9.72%、7.72%和7.22%,29年生时叶生物量所占比例分别为3.64%、2.64%和2. 03%,皮生物量所占比例分别为9.74%、7.66%和7.18%;根生物量所占比例的排列顺序是250>1 000>500株·hm-2的林分,27年生时分别为15.80%、14.36%和12.40%,2 9年生时分别为15.51%、14.34%和12.37%。在1 000株·hm-2小黑杨林分中,干生物量比例最大(27年生时为63.34%,29年生时为63.28%),叶生物量所占比例最小(27年生时为3.63%,29年生时为3.64%),说明在大密度林分中,树木自然整枝良好,干形好,干生物量所占比例大。在500和250株·hm-2的小黑杨林分中,有较大的枝生物量比例,却有较小的叶生物量比例,反映了该密度小黑杨林分自然整枝不良,叶的生长效率较低,应人工去掉下部叶较少的枝,既可以节省树木营养,又不会对叶的生长造成太大的影响,同时可以减少木材节疤,提高木材质量。
|
|

由表 3可知,3种密度27年生小黑杨生物量的径阶分布呈现出“中间高两边低"的分布规律。在1 000株·hm-2的小黑杨林分中,<18 cm的生物量占总生物量的17.26%,18~22 cm的生物量占72.37%,>22 cm的株数占总生物量的10.37%。500株·hm-2的小黑杨林分生物量主要集中在20~26 cm的径阶范围之内,占总生物量的90.02%,<20 c m树木生物量占4.39%,>26 cm㎝树木生物量占5.59%。在250株·hm-2的小黑杨林分中,生物量主要集中在径阶为26~30 cm的树木中,占总生物量的68.28%,<26 cm的树木生物量占8.89%,>30 cm㎝树木生物量占22.83%。在3种密度小黑杨林分的器官生物量分配中,根生物量所占比例随着径阶增加呈增加的趋势,其他器官生物量所占比例在不同密度林分中表现出不同的变化趋势。在1 000株·hm-2小黑杨林分中,随着径阶增加,干生物量所占比例逐渐降低,枝、叶、皮生物量比例逐渐升高,说明大密度林分有利于培育小径材;在500株·hm-2的小黑杨林分中,随着径阶增加,干、叶、皮生物量所占比例逐渐下降,而枝生物量所占比例逐渐升高,反映了该密度林分自然整枝不良,并且枝干生长的叶较少,应该及时修去下部枝条,既节省营养,又提高木材质量;在250株·hm-2林分中,随着径阶增加,树干生物量所占比例逐渐增加,枝、叶、皮生物量所占比例逐渐降低,揭示了小密度林分适合培养大径材。
|
|
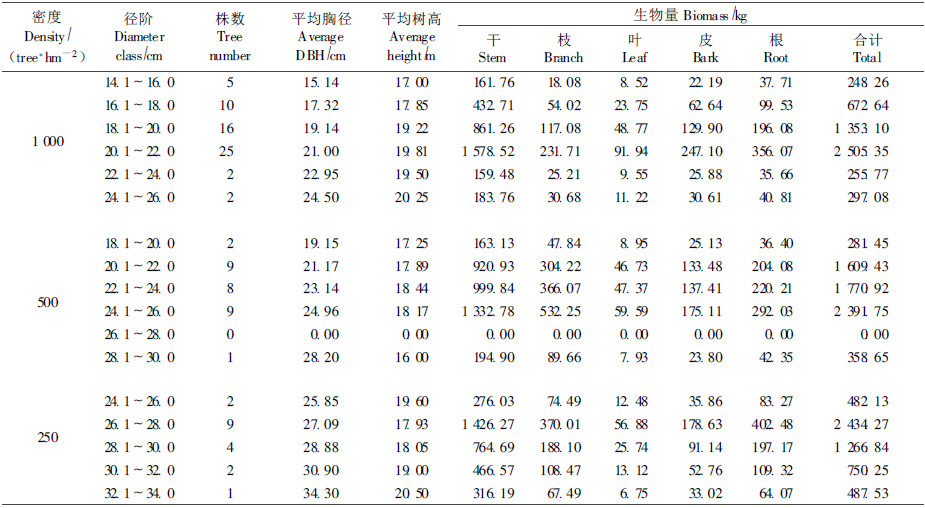
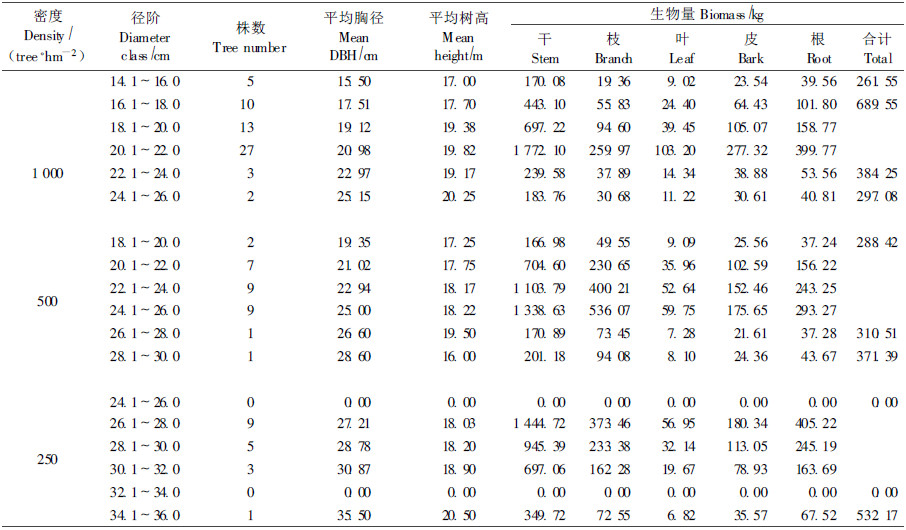
表 4列出了3种密度29年生小黑杨生物量的径阶分布,与27年生小黑杨人工林生物量的径阶分布规律相似,也呈现出“中间高两边低"的分布规律。器官生物量在绝对值上比27年生小黑杨器官生物量有一定的增加,但其在各径阶间的分配比例与27年生小黑杨人工林器官生物量的分配比例相似,说明27年生小黑杨已经进入成熟期,生长缓慢,生物量在各器官间的分配不再有大的变化。
|
|
所调查的小黑杨人工林虽为同龄林,但由于小黑杨个体的差异及其所处的环境条件不完全相同,树木的生长产生了差异,这种差异也造成了林木生物量的差异。图 1揭示了不同密度27年生和29年生小黑杨人工林株数与生物量在不同径阶间的分配规律,3种密度27年生和29生小黑杨的株数和生物量径阶分布均表现出正态分布特征。可以看出,在3种密度的林分中,29年生小黑杨林分株数的径阶分布曲线与27年生小黑杨林分株数的径阶分布曲线几乎重叠,说明3种密度的小黑杨林分在2年内的胸径变化极小,生长极其缓慢。不同密度27、29年生小黑杨林分生物量的径阶分布规律与株数的径阶分布规律类似,也呈现出正态分布格局。不同的是3种密度林分生物量分布峰值相差不大,而株数的径阶分布峰值则是1 000株·hm-2林分远远高于500和250株·hm-2的小黑杨林分,反映了在株数相等的情况下,径阶越大,林分生物量越高。由图 1也可以看出,密度为1 000和250株·hm-2的小黑杨林分径阶分布曲线较陡峭,500株·hm-2小黑杨林分径阶分布曲线较平缓,说明了500株·hm-2的小黑杨林分胸径方差较大,林分径阶分化较明显,而1 000和250株·hm-2的小黑杨林分生长较均匀。

|
图 1 不同林龄和密度小黑杨株数和生物量的径阶分配 Fig. 1 Diameter class distribution of tree number and biomass in P. × xiaohei plantation with different ages and densities |
27年生小黑杨人工林已经进入成熟期,生长极其缓慢,到29年时胸径、生物量的增长幅度很小。密度为1 000、500和250株·hm-2的27年小黑杨林分生物量分别85.23、102.60和86.74 t·hm-2;29年生的分别为88.64、104.90和90.94 t·hm-2。在生产实践中,可根据实际需要选择适宜的栽植密度和采伐时间。
林分的生物量与林分密度、年龄和生长环境密切相关。密度500株·hm-2的华北沙地小黑杨林分生物量高于密度1 000和250株·hm-2的林分,这与唐万鹏等(20 04)的研究成果相符,当林分密度从208株·hm-2增加到625株·hm-2时,南方型杨树生物量从57.6 t·hm-2增加到最高值112.87 t·hm-2,密度继续增加到1 000株·hm-2时,生物量逐渐下降到82.83 t·hm-2;曹志伟等(2006)也发现,当林分密度从570株·hm-2增加到667株·hm-2,10年生小黑杨生物量从58.45 t·hm-2降低到11.99 t·hm-2,这与本文小黑杨生物量随着密度的变化趋势相符,密度大于500株·hm-2时,生物量随着密度的增加而降低。密度对林分的影响主要通过种内、种间竞争来实现。密度不同会影响到林木的生存竞争环境,包括林木对水、肥、光等各种可利用资源的争夺,从而影响到林分生物量和生产力(Kellomaki S, 1986)。林分年龄与林分的生物量有密切关系,随着林分年龄的增大,林分生物量也增大。密度相同的小黑杨林分,29年生生物量大于27年生的生物量,丁贵杰等(2002)对马尾松人工林生物量随林龄的变化进行研究时,也揭示了相似变化规律。生长环境的差异可以造成林分生物量的较大差异,通过资料查询可知,北方地区杨树的生物量变化范围为11.99~11 6.2 t·hm-2 (崔浪军等,2003;曹志伟等,2006),而南方杨树林的生物量为57.6 ~201.93 t·hm-2(吴泽民等,2001;唐万鹏等,2004)。在密度和林龄相同的情况下,南方杨树林生物量高于北方的杨树林生物量,这主要是由气候条件差异造成的,尤其是水分,水分是杨树生长的重要限制因子(王力等,2004)。在生产实践中,可通过改变栽植密度、适当延长采伐时间和改善生长环境等措施来提高小黑杨林分单位面积的生物量。
3种密度小黑杨林分株树和生物量的径阶分配均表现为正态分布,与杨宗武等(2001)对福建柏(Fokienia hodginsii)人工林生物量分布规律的研究结果相同。在1 000株·hm-2的小黑杨林分中72.37%的生物量集中在径阶为18~22 cm的树木;500株·hm-2的小黑杨林分生物量主要集中在径阶为20 ~26 cm的树木,占总生物量的90.02%;250株·hm-2的小黑杨林分中,生物量主要集中在径阶为26~30 cm的树木,占总生物量的68.27%。可见,较低的栽植密度有利于生物量在大径阶的分布,有利于培育大径材。
密度对小黑杨器官间生物量分配有重要影响。密度不同,器官生物量所占比例不同,其中,叶和皮生物量所占比例均随着密度增加而增加,干、枝和根生物量所占比例随密度增加没有表现出明显规律性。器官生物量所占比例在同一密度不同林龄的林分中差异很小,但在同一林龄不同密度的林分中差异较大。在3种密度中,干生物量所占比例最大,叶生物量所占比例最小。
曹志伟, 戴玉玮, 张玉柱, 等. 2006. 嫩江沙地杨树用材林生产力的研究. 防护林科技, (2): 9-11. DOI:10.3969/j.issn.1005-5215.2006.02.004 |
崔浪军, 梁宗锁, 韩蕊莲, 等. 2003. 沙棘-杨树混交林生物量、林地土壤特性及其根系分布特征研究. 林业科学, 39(6): 1-7. DOI:10.3321/j.issn:1001-7488.2003.06.001 |
丁贵杰, 王鹏程. 2002. 马尾松人工林生物量及生产力变化规律研究Ⅱ. 不同林龄生物量及生产力.林业科学研究, 15(1): 54-60. |
唐守正, 张会如, 胥辉. 2000. 相容性生物量模型的建立及其估计方法. 林业科学, 36(增刊): 19-27. |
唐万鹏, 王月容, 郑兰英. 2004. 南方型杨树人工林生物量与生产力研究. 湖北林业科技, (sp1): 43-47. |
王力, 邵明安, 侯庆春, 等. 2004. 不同水肥条件对杨树生物量的影响. 西北农林科技大学学报, 32(3): 53-58. DOI:10.3321/j.issn:1671-9387.2004.03.013 |
吴泽民, 陈美工. 2001. 安徽长江滩地杨树人工林生物量和养分积累. 应用生态学报, 12(6): 806-810. DOI:10.3321/j.issn:1001-9332.2001.06.002 |
杨宗武, 谭芳林, 肖祥希, 等. 2001. 福建柏人工林生物量的研究. 林业科学, 36(专刊1): 120-124. |
赵天锡, 陈章水. 1994. 中国杨树集约栽培. 北京: 中国科学技术出版社.
|
张会儒, 赵有贤, 王学力, 等. 1999. 应用线性联立方程组方法建立相容性生物量模型. 林业资源管理, 6: 63-67. |
朱春全, 刘晓东, 张启, 等. 1997. 集约与粗放经营杨树人工林生物量的研究. 东北林业大学学报, 25(5): 53-56. |
Anna A S, Diane L M, Steven A. 2000. Competition between native Populus deltoids and invasive Tamarix ramosissima and the implications for reestablishing flooding disturbance. Conservation Biology, 14(6): 1744-1754. DOI:10.1111/j.1523-1739.2000.99306.x |
Baldwin J R, Burkhart H E, Westfall J M, et al. 2001. Linking growth and yield and process models to estimate impact of environmental changes on growth of loblolly pine. Forest Science, 47(1): 77-82. |
Gower S T, Vogel J G, Norman J M, et al. 1997. Carbon distribution and aboveground net primary production in aspen, jack pine, and black spruce stands in Saskatchewan and Manitoba, Canada. Journal of Geophysical Research, 102(D24): 29-41. |
Houghtou J T, Ding Y, Griggs D J, et al. 2001. Climate change 2001 in the Scientific Basis. Cambridge: Cambridge University Press.
|
Jorg K, Ilka H, Carsten M, et al. 2003. Elevated pCO2 affects N-metabolism of young poplar plants (Populus tremula×P. alba) differently at deficient and sufficient N-supply. New Phytologist, 157: 68-81. |
Kellomaki S. 1986. A model for the relationship between branch number and biomass in Pinus sylvestris crowns and the effect of crown shape and stand density on branch and stem biomass. Scandinavian Journal of Forest Research, 1: 455-472. DOI:10.1080/02827588609382437 |
Kurz W A, Apps M J. 1999. A 70-year retrospective analysis of carbon fluxes in the Canadian forest sector. Ecological Applications, 9(2): 526-547. DOI:10.1890/1051-0761(1999)009[0526:AYRAOC]2.0.CO;2 |
Liu J, Chen J M, Cihlar J, et al. 1999. Net primary productivity distribution in the BOREAS study region from a process model using satellite and surface data. Journal of Geophysical Research, 104(D22): 735-354. |
Long S P, Ainsworth E A, Rogers A, et al. 2004. Rising atmospheric carbon dioxide:Plant FACE the future. Annual Review of Plant Biology, 55: 591-628. DOI:10.1146/annurev.arplant.55.031903.141610 |
Norby R J, Wullschleger S D, Gunderson C A, et al. 1999. Tree responses to rising CO2 in field experiments:implications for the future forest. Plant, Cell & Environment, 22: 683-714. |
Victoria E W, Carl J B, Xin-Guang Zhu, et al. 2005. Gross primary production is stimulated for tree Populus species grown under free-air CO2 enrichment from planting through canopy closure. Global Change Biology, 11: 644-656. DOI:10.1111/j.1365-2486.2005.00934.x |