 2007, Vol. 43
2007, Vol. 43文章信息
- 王鸿斌, 张真, 孔祥波, 刘随存, 沈佐锐.
- Wang Hongbin, Zhang Zhen, Kong Xiangbo, Liu Suicun, Shen Zuorui.
- 入侵害虫红脂大小蠹的适生区和适生寄主分析
- Preliminary Deduction of Potential Distribution and Alternative Hosts of Invasive Pest, Dendroctonus valens (Coleoptera: Scolytidae)
- 林业科学, 2007, 43(10): 71-76.
- Scientia Silvae Sinicae, 2007, 43(10): 71-76.
-
文章历史
- 收稿日期:2006-03-09
-
作者相关文章


2. 山西省林业科学研究院 太原 030012;
3. 中国农业大学农业与生物技术学院IPMIST实验室 北京 100094
2. Shanxi Academy of Forestry Taiyuan 030012;
3. IPMIST Lab., College of Agriculture and Biotechnology, Chinese Agricultural University Beijing 100094
防止外来有害生物造成更大危害的重要手段之一是阻止其进入适合生存的地区(Tucker et al., 1995; Richard et al., 1997)。研究传入物种适生区和适生寄主对于入侵害虫的管理和决策有很大意义。对于入侵害虫适生区域的研究,由于入侵害虫的高风险性,基本不允许进行野外试验。一般通过气候和生物地理条件来分析预测入侵物种的适生区域,随计算机技术的进步和推广应用,适生区研究的手段也在不断更新和提高(刘红霞等,2001)。不仅统计模型应用于适生区分析,还产生了一些生物气候分析软件如CLIMEX、BIOCLIM、GARP等,近年更将地理信息系统结合到分析中去(徐汝梅,2003)。对于入侵害虫适生寄主的研究目前主要依赖于原产地的寄主资料来进行分析判断,并结合实际入侵发生区内潜在寄主接种害虫后的生物学观察来进行。但由于试验条件的局限性,多数是人为对害虫进行取食选择试验,试验周期长,受影响因素多,也难以扩展到整个评价区域,很难反应实际的潜在适生寄主状况,一般只是定性地评价其能够入侵与否。
被定为入侵害虫的红脂大小蠹(Dendroctonus valens),是1998年在我国山西沁水被首次发现的。近年来随着森保管理部门的调查跟踪,发现该虫已经在山西省10市61县、河北省5市17县、河南省9市21县、陕西省2市3县形成广泛种群定殖,其寄主除了当地的油松(Pinus tabulaeformis)外,还有少量当地的白皮松(Pinus bungeana)等其他针叶树种(苗振旺等,2001;王鸿斌等,2005;张历燕等,2002)。根据该虫在北美发生的经纬度和寄主情况,其分布区应该覆盖我国绝大部分区域,其寄主也要涉及我国绝大多数针叶树种(宋玉双等,2000)。而事实上由于入侵种本身种群来源的差异性,其存在着对我国环境的适应性问题。本研究将根据红脂大小蠹种群的暴发和扩散记录,结合其基本生物学特性以及暴发前后发生区气候的变化来比较分析该虫在我国的适生区域,并尝试根据该害虫对于寄主植物的取食选择趋性的主要化学信息物质的相似性来判定其潜在适生寄主。
1 材料与方法 1.1 红脂大小蠹历年分布扩散情况目前发生存在区和暴发记录的分析是进行潜在适生区推测的首要条件。根据国家林业局森林病虫害防治总站提供的历史发生数据,应用地理信息系统软件Arcview3.2,做出红脂大小蠹自发生以来在我国的发生及蔓延状况的扩展动态图。
1.2 气候适生区图一定时期内稳定种群的突然暴发与气候相关性较强,为了解在红脂大小蠹暴发时起关键作用的是何种气象因子,笔者从国家气象局购进了相关年代的气象资料,包括全国305个气象观测站1980年1月至1997年12月的逐月气象观测资料,项目有:平均气温、最高气温、最低气温、降水量、日照时数、相对湿度、蒸发量和辐射量。
根据国家气象局气象站的历史气候资料,结合目前发生区的地理信息系统资料,分析红脂大小蠹发生区内相关站点的气候相似性的主要作用因子。根据这些因子的取值范围用Arcv iew3.2软件做出各个因子相似条件的分布图,最后不同因子叠加得到该虫的气候适生区图,并进行分析。
1.3 寄主适生范围分析在北美,红脂大小蠹的针叶寄主树木大于40余种(Smith,1961;苗振旺等,2001;宋玉双等,2000)。我国幅员辽阔,针叶树种资源也比较丰富,不可能在所有地方的潜在寄生树种上进行接种试验,确定入侵可能性,移植或砍木段的方法由于要砍比较大的活立木也不可取。根据红脂大小蠹对于寄主植物的选择趋性是由特定的化学信息物质决定的特点,探索根据对不同针叶树挥发性成分的分析判断比较,确立潜在的寄主树种,并根据寄主树种的分布区域和范围确定该虫的潜在寄主范围。
2 结果与分析 2.1 红脂大小蠹分布扩散趋势红脂大小蠹是1998年在山西省沁水、阳城开始发现为害的,图 1a为目前我国红脂大小蠹发生的4个省区图,中间的阴影区域则为沁水和阳城。图 1b是2001年山西省发生的市县分布状况,图 1c则显示了2003年在山西、河北、河南、陕西发生的市县分布状况。

|
图 1 红脂大小蠹在我国发生扩散情况(1998年,2001年,2003年) Fig. 1 Spread of the D. valens in China in 1998, 2001, 2003 |
在虫害开始阶段,由于对该虫的分布和生物学特性不甚了解,生产管理部门没有找到相应的防治办法,遇到虫害致死木基本上都整株伐除,对伐后的树桩残根没有进行相应的处理。经过调查,绝大部分濒死树上红脂大小蠹入侵产卵并孵化为幼虫后,都向树根纵深取食,有的达到2~3 m深,并在那里化蛹成室。至成虫扬飞期才不等时地从根部蛀出羽化孔钻出地面。因此仅伐除危害树地上部分实际上未能抑制或降低虫口的密度,反而使地温由于没有树荫的遮蔽而升高,缩短了该虫的生活历期,加速了羽化扬飞且使之更易向四周扩散。因此在开始的几年其发生蔓延的范围非常迅速,发生面积呈几何级数增长。
近2年来,通过对该害虫各项研究的深入,过去的砍伐濒死树的行为已经基本不再采用,加之引诱剂监测等监测治理措施的推广应用,其扩散的速度和种群数量已经明显呈连年下降趋势。
2.2 气候适生区红脂大小蠹目前普遍被认为是20世纪80年代由北美洲传入的(苗振旺等,2001),虽然其具体时间还尚未证实,但目前通过对在我国和北美各地所采集的不同活体昆虫样本进行的分子遗传学分析证实,其与美国本地种的亲缘关系很近。而该传入害虫大概经过十几年的种群潜伏定殖期后,在20世纪末才得以种群暴发。
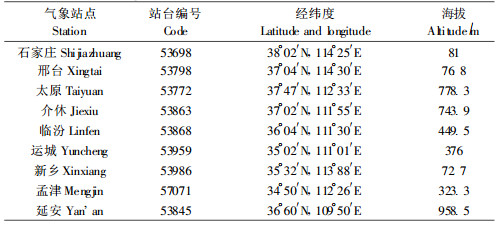
305个站点的位置在我国的分布见图 2,将图与该虫目前分布区图叠加可知在发生区及附近范围内的气象站点为9个(表 1)。这9个气象台站平均分布于目前红脂大小蠹的实际发生区,这些台站的气候观测数据基本可反应该区域大范围的气候特征状况。由于1998年该虫开始大规模发生,故选择发生前1年1 997年的气象资料数据进行分析,以确定何种气象因素在害虫发生时起决定作用。

|
图 2 用以分析害虫潜在分布区的气象站点的分布示意 Fig. 2 Sketch for distribution of the meteorological station over the country used in prediction |
|
|
分别将这9个台站的1997年度每月的气候观测数据从数据库中提出来,加入附近其他相近无害虫发生的6台站相应的数据(包括西安、郑州、五台、安阳、卢氏、榆林)。以有害虫发生为1、无害虫发生为0建立2组总体数据,即9组发生为1,另外无发生为0。根据这2组分类已定结果, 对1997年每月平均温度(WD9701—WD9712)、每月平均相对湿度(SD9701—S D9712)、每月总降水(JS9701—JS9712)、每月平均日照时间(RZ9701—RZ9712)总计48个变量利用多元统计软件Spss11.0进行判别分析,确定对分类结果贡献最大的因子。
在Spss11.0判别统计分析中,统计方法采用向前逐步判别分析,F值大于3.84(P<0.0 5)则进入判别模型,小于2.71(P>0.10)则不能进入下一轮筛选。
经过最终的判别选择,只有1个参数被模型保留作为分组的判别特征,即SD9702,F值为6.117。而以此单一特征对原有15组数据进行分类判别,准确率为73.3%;根据其他14组数据的结果判定另外一组数据的分析准确率也为73.3%。由此可见1997年春季湿度因子在害虫发生上起了关键作用,这也与实际情况非常吻合。1997年春季地面土壤和林间干燥,空气湿度极端低,非常有利于红脂大小蠹的越冬幼虫和成虫越冬种群的大量存活并羽化,造成种群暴发。
除了此判别特征外,其他因子中对分类组别贡献最大的因子为JS9704,虽然没有被判别模型保留为特征指标,但在剔除了春季湿度的因素SD9702后,以同样参数方法向前逐步回归也成为唯一的保留特征。这说明春季降水在抑制红脂大小蠹种群发生中也起到重要作用。
综合1998年害虫发生的历史事实和特别干旱的历史气象条件,可以推断,1997年林间的干旱气候和春季少雨的条件为红脂大小蠹种群增长提供了非常有利的环境因素,从而造成该虫在1998年的大规模发生。
根据得到的1997年2月的湿度和1997年4月降水为红脂大小蠹发生的关键决定因子,以此年的平均湿度(55%~66%)和总降水数值(242~423 mm)为取值范围,分别在全国气候平均湿度和降水图上预测该虫的气候适应分布,得到结果见图 3和图 4。

|
图 3 以湿度为参数进行的适生区预测示意 Fig. 3 Sketch for potential distribution based on humidity |

|
图 4 以降水为参数进行的适生区预测示意 Fig. 4 Sketch for potential distribution based on precipitation |
图 3、4结果与目前发生区图比较可得出以下2点结论:1)这2因子的单独预测结果基本相互重叠;2)这2图预测的适宜发生区明显在目前发生区之外,而且为更加干旱地区。
虽然目前由于寄主树种和虫源问题等其他相关原因,在该气候适生区域内仅有少量和局部发现危害,但由于这一区域具有极为适合红脂大小蠹暴发的气候因素,如果这种害虫一旦传入则对其内相关树种危害将更大。而相对于目前发生区而言,由于气候条件的改善和预测治理手段的进步,如果没有进一步的灾害天气,该虫将不会有太大的灾害发生。
2.3 寄主适生范围寄主植物的挥发物成分在红脂大小蠹选择入侵寄主行为时起到非常重要的作用,前人对分布于我国各地的不同针叶树种的精油或树干树叶的挥发物很多进行过相关研究(金幼菊等,1994;侯冬岩等,1999;赵成华等,1995)。以红脂大小蠹目前寄主挥发物油松的主要萜烯类成分为对照指标(王鸿斌等,2005;Sun et al., 2004), 根据其他地区针叶寄主植物挥发物的相似性判断寄主是否作为红脂大小蠹的潜在寄主,然后根据该寄主的分布情况作出潜在的发生区预测。
红脂大小蠹目前发生区内的针叶树种除油松外,还有白皮松、白杄 (Picea meyeri)、华北落叶松(Larix principis-rupprechtii)、臭冷杉(Abies nephrolepis)等针叶树种,除了油松大量受害外,发现少量白皮松和白杄也受到了侵害。
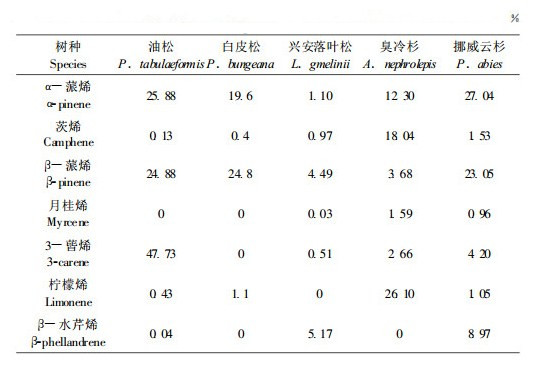
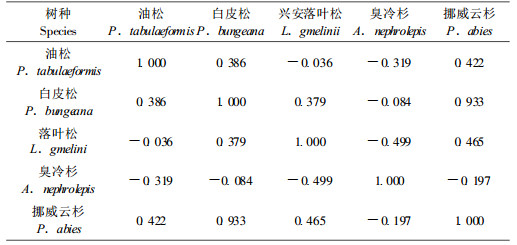
表 2是前人所测各种针叶树种的萜烯类挥发物组成的相对含量(王鸿斌等,2005;宋湛谦等,1998;严善春等,1999;徐永红等,1994;陈鹏等,2001)。在这些记录中,有些并不是红脂大小蠹发生区当地树种,但是都分别与当地种有很近的亲缘关系。从萜烯成分看,虽然其成分比例与油松有很大不同,但都含有对红脂大小蠹起引诱作用的活性成分。表 2进行相关性分析得到了这几种针叶树种在萜烯挥发物成分与油松相似关系矩阵表(表 3)。从挥发物组成比例相似性分析,白皮松和挪威云杉(P. abies)最为相似,相似系数0.933, 而与油松关系稍近的也为这两者。白皮松和与挪威云杉亲缘较近的白杄正是红脂大小蠹发生区内除油松外的另外受害针叶树种。因此在虫害潜在分布区内只要气候适宜,这类在挥发物类似于油松的针叶树种被侵害的可能性非常大。
|
|
|
|
另一方面,由于红脂大小蠹的生物学习性,也可能对不同寄主树种的挥发物产生地区适应性。如同油松在美洲原产地并无分布而在我国则成为主要寄主一样,在发生区内相关的其他针叶树种随着害虫的适应性变异也有相当大的被侵害风险。根据我国主要针叶树种的地区分布状况,排除人为传播因素,距离红脂大小蠹目前分布越近的针叶林其潜在受害的可能性越高。除了已广泛受害的油松属于高风险树种外,以下分布居于或邻近目前危害区的针叶树种具有潜在风险,松属的白皮松、华山松(P.armandii)、赤松(P. densiflora)、黑松(P. thunbergi);落叶松属的兴安落叶松、华北落叶松; 云杉属的云杉(P. asperata)、白杄、青杄 (P. wilsonii)、大果青杄 (P. neoveitchii);冷杉属的臭冷杉。
3 结论与讨论红脂大小蠹在1998年突然发生的原因与其前1年春季的气候干旱、降水偏少以及林间湿度相关。正是由于这样的气候条件造成寄主树木表皮干燥,害虫侵入成功的几率增加并大量产卵,使幼虫种群数量增加,造成第2年油松大量死亡。
根据1997年春季发生区的湿度和降水因子的取值范围来预测的红脂大小蠹气候适宜区,明显与现在实际发生区不符合,说明现发生区并不是最佳的气候适宜区,随着气候的好转该虫危害会逐渐下降。而所预测产生的适生气候区如果存在适宜寄主,一旦传入该虫将会严重发生。
红脂大小蠹的发生状况由于缺乏实际的监测数据,只能以行政区域为对象作图,也没有实际发生程度和种群数量的差别。而实际上更为精确的地理信息系统图都应当反应在各个林场林班的林相图中,这还有待于生产管理部门提高监测手段和水平,提供更加确凿的数据,以便更好地应用服务于生产管理。
本研究以中央气象台站的数据来分析红脂大小蠹发生区的情况,而这些台站在害虫发生区不仅偏少,而且这些气象站点的实际位置与红脂大小蠹实际发生地还有相当的距离,如新乡市、邢台市和石家庄市,其气象站的位置海拔明显不同于红脂大小蠹的发生区。由于该虫基本发生在以油松为主要树种的山区,都是位于这些城市的临近山区县,其小气候也有所不同。这可能是造成在判别分析中误判结果的重要原因。如果能够以更精确的小环境尺度来记录观测红脂大小蠹各个发生区的气象状况,无疑会使其精度提高和更有说服力。
以寄主挥发物的相似性来分析白皮松和云杉的挥发性成分与油松最为接近,又处在发生区,是较高风险的潜在寄主树种类,但有关所有寄主树种的分布仅有省区行政的分布,缺乏地理信息系统的数据没有能与气候适宜区进行叠加,在今后的研究中与之结合才更有说服力。
陈鹏, 赵涛, 李丽莎. 2001. 挪威云杉幼树韧皮部挥发性物质的测定. 云南林业科技, (2): 58-60. DOI:10.3969/j.issn.1672-8246.2001.02.013 |
侯冬岩, 张维华, 回瑞华. 1999. 松属植物挥发性成分分析——辽宁不同地区的油松松针挥发油成分的研究. 鞍山师范学院学报, 1(3): 78-84. |
金幼菊, 吴京科, 孙福, 等. 1994. 油松针叶精油萜烯组成的研究(Ⅰ)——油松针叶的萜烯组成及其与其它两针松的比较. 北京林业大学学报, 16(3): 38-47. |
刘红霞, 温俊宝, 骆有庆, 等. 2001. 森林有害生物风险分析研究进展. 北京林业大学学报, 23(6): 46-51. DOI:10.3321/j.issn:1000-1522.2001.06.011 |
苗振旺, 周维民, 霍履远, 等. 2001. 强大小蠹生物学特性研究. 山西林业科技, (1): 34-40. DOI:10.3969/j.issn.1007-726X.2001.01.010 |
宋玉双, 杨安龙, 何嫩江. 2000. 森林有害生物红脂大小蠹的危险性分析. 森林病虫通讯, (6): 34-37. |
宋湛谦, 梁志勤, 刘星. 1998. 白皮松松脂的化学特征及其分类学意义. 植物分类学报, 36(6): 511-514. |
王鸿斌, 张真, 孔祥波, 等. 2005. 油松萜烯类挥发物释放规律与红脂大小蠹危害关系研究. 北京林业大学学报:自然科学版, 27(2): 75-80. |
徐汝梅. 2003. 生物入侵数据集成、数量分析与预警. 北京: 科学出版社.
|
徐永红, 申仁花, 李东浩, 等. 1994. 延边地区臭冷杉精油化学成分的研究. 延边大学学报:自然科学版, 20(4): 39-42. |
严善春, 胡隐月, 孙江华, 等. 1998. 落叶松挥发性物质与球果花蝇危害的关系. 林业科学, 35(3): 58-62. |
张历燕, 陈庆昌, 张小波. 2002. 红脂大小蠹形态学特征及生物学特性研究. 林业科学, 38(4): 95-99. DOI:10.3321/j.issn:1001-7488.2002.04.016 |
赵成华, 伍德明, 阎云花, 等. 1995. 马尾松针叶挥发性成分的鉴定及其对马尾松毛虫的触角电位反应. 林业科学, 31(2): 125-131. |
Richard S H, Hamilton. 1997. Predicting invasion of woody plants introduction in North America. Conserv Bio, 11: 193-203. DOI:10.1046/j.1523-1739.1997.95473.x |
Smith R H. 1961. Red turpentine beetle. Washington D C: U. S: Department of Agriculture, Forest Service.
|
Sun J H, Miao Z, Zhang Z, et al. 2004. Red Turpentine Beetle, Dendroctonus valens LeConte (Coleoptera: Scolytidae), response to host semiochemicals in China. Environmental Entomology, 33(2): 206-212. DOI:10.1603/0046-225X-33.2.206 |
Tucker K C, Richardson D M. 1995. An expert system for screening potentially invasive alien plants in south Africa fynbos. J Environ Manag, 44: 309-338. DOI:10.1016/S0301-4797(95)90347-X |