 2007, Vol. 43
2007, Vol. 43文章信息
- 牟长城, 孙晓新, 倪志英, 杨明, 张娜.
- Mu Changcheng, Sun Xiaoxin, Ni Zhiying, Yang Ming, Zhang Na.
- 大兴安岭林区沼泽-森林人工造林效果综合评估
- Comprehensive Evaluation of the Effects Planting in Swamp-Forest in Daxing'an Mountains
- 林业科学, 2007, 43(10): 51-58.
- Scientia Silvae Sinicae, 2007, 43(10): 51-58.
-
文章历史
- 收稿日期:2006-01-12
-
作者相关文章


湿地森林中蕴藏着丰富植物资源、泥炭资源(王荣芬,1994),又是野生动植物重要生境(Ohlson et al., 1997;郎惠卿等,1999),同时具有涵养水源、调节河川径流与区域气候、净化水质等多种生态功能,对保持生态平衡和物种多样性具有重要意义(马学慧等,1991;郎惠卿等,1999)。因此,目前全球对这类土地的保护和合理利用问题十分关注。
国外有关湿地森林研究主要集中在湿地森林群落组成结构、演替规律、生物多样性、生产力及退化植被恢复等方面。如欧洲和北美东部温带泛滥平原森林的演替研究结果表明:在洪水侵蚀和堆积的生境上首先是由杨属(Populus)、柳属(Salix)和赤杨属(Alnus)的一些耐水湿树种形成先锋群落,经过几十至上百年时间的发育,逐渐被栎树(Quercus)、榆树(Ulmus)、水曲柳(Fraxinus)和槭树(Acer)等组成的硬阔叶混交林所取代(Barnes,1985;Schnitzler,1995a;1995b),且因水淹程度及微地形变化常形成不同的森林群落类型(Sharitz et al., 1993;Hodges,1995)。这一演替规律应是温带泛滥平原森林经营的重要依据。温带泛滥平原森林较其邻近的上部森林具有更高的物种丰富度(Hodges,1995;Schnitzler et al., 2005),温带湿地森林也具有较高的生产力(Schnitzler,1994;Kellison et al., 1998)。因此,温带湿地森林在生物多样性维持及促进区域经济发展方面具有重要作用。但由于过渡利用及不合理经营,使得这一宝贵资源遭到了严重的破坏,如美国密西西比河流域因发展农业已使早期的1 000万hm2湿地森林减少到200万hm2(MacDonald et al., 1979),但其中大部分土地由于经常遭受春末夏初洪水危害,不适宜发展农业,美国农业部正在实施自然保护区计划与湿地保护区计划恢复湿地森林(Shepard, 1995;Stanturf et al., 1998),在湿地森林恢复实践中特别重视泛滥平原微生境的变化及造林树种对生境的要求,认为地势的微小变化将导致生境质量和树种适应性的较大变化(Hodges,1997;1998;Stanturf et al., 2001)。
东北林区是我国湿地森林的主要分布区(郎惠卿等,1999),在维持区域生态平衡和生物多样性保护方面具有重要作用。目前有关东北湿地森林的研究工作主要集中在湿地森林类型与分布、形成与演替、保护与利用等方面(郎惠卿,1996;郎惠卿等,1999;牟长城等,1998; 2001a; 2001b; 2004;牟长城,2003)。而对大、小兴安岭湿地的改造利用是学术界争论的焦点问题之一,即湿地是否应进行排水造林或排水提高林分生产力,有些学者认为湿地在维护该地区生态环境和景观完整性以及在生物多样性保护等方面均有不可替代的重要作用,对其应以保护和保护性利用为主,不宜大规模改造造林(徐化成,2004);也有学者认为湿地排水改造后能够改善生境条件促进林木生长,对木材生产具有一定的积极意义(李博生等,1996)。因此,探讨湿地生态保护与木材生产相协调的可持续经营途径就显得尤为重要。但目前相关研究工作主要涉及到湿地资源的合理利用、排水改造对植被组成结构与动态、土壤养分循环、建群种的生长过程等方面的影响研究,如季节性积水型湿地适宜改造造林(张武文等,1995b),湿地经过排水改造或造林后湿生植物类群将逐渐被中生植物类群所取代,湿地植物群落逐步向高地森林群落发展(马学慧等,1991;张武文等,1996),土壤的地表形态、剖面构造、水热状况、理化性质及土壤肥力等有不同程度改善(张武文等,1995a;1996;白燕等,1996;许红梅等,2000),并促进了树木的生长(李博生等,1996)。总之,目前尚缺乏对湿地人工林生产力与生物多样性的系统研究,也没有考虑水分环境梯度的影响以及与相应生境地段天然湿地森林的对比分析,因此难以客观地评价湿地人工林的整体功能与价值。
本文以大兴安岭22年生人工兴安落叶松(Larix gmelinii)沼泽林为研究对象,采用样带调查和对比分析方法,研究人工兴安落叶松沼泽群落的组成结构、生产力、生物多样性沿沼泽-森林过渡带环境梯度的变化规律,并与其相应生境地段的天然沼泽林群落进行对比分析,综合定量评价沼泽-森林人工造林的效果,探讨沼泽-森林有效恢复与合理利用途径,以便为科学地经营管理大兴安岭湿地森林资源提供依据。
1 研究区概况与研究方法 1.1 研究地概况研究地点位于大兴安岭东南部松岭林业局(50°09′—51°24′ N,123°29′—125°50′ E)。本区气候属于寒温带大陆季风气候,冬季寒冷而漫长;夏季温暖,但时间较短,最长不超过1个月,年平均气温-3 ℃,≥10 ℃的积温约为1 600 ℃。年平均降水量600 mm,6— 8月的降水量常占全年降水量的60%以上。本区的土壤主要为棕色针叶林土、暗棕壤、灰色森林土,低洼地带还有成片隐域性的草甸土和沼泽土。土壤永冻层在部分地区呈岛状分布,而季节冻层则在全区普遍存在。地带性植被为寒温带针叶林。在低洼地带分布有沼泽湿地及森林与湿地的过渡类型,如沿着沼泽至森林方向过渡带环境梯度依次分布有典型草本沼泽、毛赤杨(Alnus sibirica var. hirsuta)沼泽林、白桦(Betula platyphylla)沼泽林和兴安落叶松沼泽林。20世纪80年代初对沼泽-森林过渡区域进行过人工造林改造,对改造地排水和机械整地,然后人工栽植了兴安落叶松,初植密度为3 000株·hm-2;2005年对林分进行了调查,林龄为22年,保存率40% ~60%,郁闭度为0.9。由于微地势的存在,导致过渡带各部分受水淹的程度和周期各不相同,从典型草本沼泽到高地森林,随着地势的升高,水淹周期逐渐缩短,受水淹的程度逐渐减轻,因而形成了过渡带上从典型草本沼泽到高地森林的水分环境梯度。由于水分梯度与标准地在过渡带上的距离相对应,为表述方便,这里以标准地在过渡带上的距离代替,下同。林分平均树高和平均胸径沿沼泽到森林的环境梯度分别为:人工落叶松沼泽林3.25 m、4.67 c m (0~100 m),7.82 m、9.90 cm(100~200 m),9.10 m、11.82 cm(200~300 m),9.23 m、13.84 cm(300~400 m);对照地天然沼泽林平均树高和平均胸径沿沼泽到森林的环境梯度为:0~100 m无乔木,其他各地段分别为8.29 m、5.27 cm(100~200 m),12.20 m、8.17 cm(200~300 m),9.12 m、4.51 cm(300~400 m)。林下植被较稀疏,灌木有柳叶绣线菊(Spiraea salicifolia)、笃斯越橘(Vaccinium uliginosum)、柴桦(Betula fruticosa)和沼柳(Salix rosmarinifolia var. brachypoda),不能成层;草本层可见膨囊苔草(Carex schmidtii)、小叶章(Calamagrostis angustifolia)、小白花地榆(Sanguisorba parviflora)、龙江凤毛菊(Saussurea amurensis)等。
1.2 研究方法 1.2.1 调查方法2005年7—8月对研究地进行调查,沿着从沼泽到森林方向的过渡带环境梯度,在人工落叶松沼泽林和天然沼泽林内分别设置一条宽20 m、长400 m的调查样带,在各样带中每间隔1 00 m,分别设置一块20 m×30 m的标准地,共设置8块。然后,将每块标准地划分为24个5 m ×5 m的样方,在样方内进行每木调查,测定乔木种类、数量、年龄(生长锥法)及胸径生长量,分析蓄积生长量及乔木物种多样性;在各标准地内随机设置5个5 m×5 m的样方,调查灌木种类及个体数量,分析灌木物种多样性;在各标准地随机设置10个1 m×1 m的小样方,调查草本植物种类及其个体数量,分析草本物种多样性。在每个标准地内,选取一个典型地点,挖土壤剖面,记述土壤剖面结构特征,测量泥炭层厚度及地下水位深度,并结合现有人工沼泽林的养分影响研究结果,分析人工沼泽林的环境影响效果,从而揭示人工兴安落叶松沼泽林群落的组成结构、蓄积生长量、生物多样性沿沼泽-森林过渡带环境梯度的变化规律,并与其相应生境地段的天然沼泽森林群落进行对比分析,综合定量评价人工造林效果,探讨沼泽-森林保护与合理利用途径。
1.2.2 分析方法1) 林分蓄积量的计算方法:利用林业部大兴安岭林管局局标准(一元材积表);2)物种丰富度S=在样地内出现的物种数;3) Shannon-Wiener多样性指数: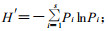
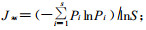
大兴安岭人工落叶松沼泽林与天然沼泽林群落建群种沿沼泽-森林过渡带环境梯度的更替规律不同(表 1)。天然沼泽林群落组成和层次结构一般均比较复杂,除典型草本沼泽外,均具有明显的乔木、灌木和草本3层垂直结构,沿着沼泽到森林方向的过渡带环境梯度,依次出现有典型草本沼泽、毛赤杨沼泽(6毛3白1钻)、白桦沼泽(5毛4白1落)和落叶松苔草沼泽(10落),其乔木层由毛赤杨、白桦、兴安落叶松依照不同比例混交成林,且群落建群种呈现出明显的更替现象;灌木层发达,在林下形成以柴桦、柳叶绣线菊、笃斯越橘和沼柳为主的灌木层,盖度约为50%~80%;草本层较发达,形成以大叶章(Calamagrostis laugsdorffii)和丛苔草(Carex caespitosa)为优势种的草本层,伴生种有兴安鹿药(Smilacina dahurica)、七瓣莲(Trientalis europaea)、龙江凤毛菊等,盖度为70%左右。而人工落叶松沼泽林群落组成结构比较单一,垂直层次结构比较简单。乔木层树种较少,沿着沼泽到森林方向的过渡带环境梯度,每间隔100 m,群落树种组成结构分别为10落、6落2白2钻、10落和10落,均以落叶松为单优种,仅在个别生境地段混有少量白桦和钻天柳(Chosenia arbitifolia),且分布不均匀;灌木发育不良,不能单独成层,仅见稀疏丛状或单株分布的柳叶绣线菊、笃斯越橘、柴桦和沼柳,盖度不足10%;草本层稀疏分布一些膨囊苔草、小叶章、地榆、龙江凤毛菊等,盖度约为20%~30%,没有明显的3层垂直结构。由此可见,人工落叶松沼泽林的乔木层、灌木层和草本层虽然仍是以湿生植物类群为主体,但各物种的组成比例和分布状况却与天然沼泽林存在较大的差异。
|
|
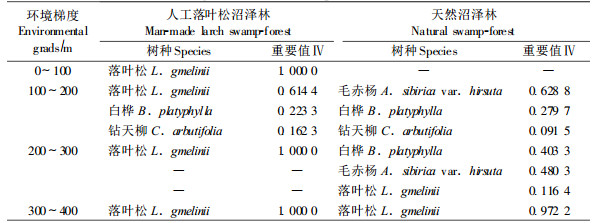
大兴安岭人工落叶松沼泽林的径级分布沿沼泽-森林过渡带环境梯度呈现规律性变化(图 1)。处于沼泽-森林过渡带0~100 m生境的人工落叶松沼泽林,其径级分布幅度为2~8 cm;随着水淹程度的降低,各地段上人工落叶松沼泽林的径级分布幅度逐步扩大到4~24 cm,且达到12 cm径级(中径材标准)以上的林木所占比例逐渐由32.7%增大到65.5%和80.7%。可见,沼泽-森林过渡带环境梯度制约着落叶松林分的生长状况,其原因主要是由于过渡带地势逐渐升高,使季节性积水周期及积水量均发生梯度变化,树木适应水分梯度变化导致生长差异。因此,培育湿地落叶松林应以该类型过渡带中上部生境地段较为适宜。

|
图 1 大兴安岭人工落叶松沼泽林沿沼泽-森林过渡生境的径级分布 Fig. 1 Diameter classes distribution of man-made larch swamp-forest along with swamp-forest ecotone in Daxing'an mountains |
大兴安岭人工落叶松沼泽林蓄积生长量沿沼泽至森林方向过渡带环境梯度呈现递增趋势(图 2)。处于沼泽-森林过渡带0~400 m各个地段的落叶松人工林,蓄积量分别为2.261 7、95.393 3、149.316 7和153.791 7 m3·hm-2。年平均蓄积生长量分别为0.10 2 8、4.349 7、6.787 1和6.990 5 m3·hm-2a-1。由此可见,沼泽-森林过渡带环境梯度对人工落叶松沼泽林蓄积生长量具有重要的影响,在沼泽-森林过渡带环境梯度不同生境地段排水造林效果差异较大,在典型沼泽生境地段排水造林(过渡带0~100 m),林分生产力极低,而在沼泽-森林过渡带中进行排水造林,可获得较高的林分蓄积生长量,特别是在过渡带的中上部生境地段上,其年蓄积生长量在6.79~6.99 m3·hm-2a-1之间,其值为黑龙江省森林年平均蓄积生长量(2.49 m3·hm-2a-1)(李坚,1994)的2.73~2.81倍,因此,该区域是具有高生产力的林业用地。

|
图 2 大兴安岭不同生境人工落叶松沼泽林和天然沼泽林林分蓄积量(A)和年蓄积生长量(B) Fig. 2 Stand volume(A) and mean annual stand volume(B) of man-made larch swamp-forest and natural swamp-forest in different habitats in Daxing'an mountains |
进一步将大兴安岭人工落叶松沼泽林与其相应生境地段的天然沼泽林的蓄积生长量对比分析得到,两者蓄积生长量沿着沼泽-森林方向的环境梯度变化趋势不同,前者呈现递增趋势,而后者则呈递减趋势。在100~200 m生境地段,毛赤杨沼泽林(林龄22年)蓄积量达到了147.205 1 m3·hm-2,年平均蓄积生长量高达6.689 8 m3·hm-2a-1,为其相应地段人工落叶松沼泽林年均蓄积生长量的153.8%;在200~300 m生境地段,白桦沼泽林(林龄50年)蓄积量为129.328 6 m3·hm-2,年均蓄积生长量为2.586 6 m3·hm-2 a-1,仅为其相应地段人工林的38.1%;在300~400 m生境地段,兴安落叶松沼泽林(林龄45年)的蓄积量为63.530 1 m3·hm-2,年均蓄积生长量为1.411 8 m3 ·hm-2a-1,仅为其相应地段人工林的20.2%。由此可见,在沼泽-森林过渡带环境梯度的下部生境地段天然毛赤杨沼泽林年蓄积生长量远高于人工落叶松沼泽林,而在其中上部生境地段,人工落叶松沼泽林的年蓄积生长量却远高于天然白桦沼泽林和兴安落叶松沼泽林。因此,沼泽-森林过渡带环境梯度对其年蓄积生长量具有重要的影响,且其对天然沼泽森林年蓄积生长量与人工落叶松沼泽林年蓄积生长量的影响机制正好相反。主要是由于不同树种的生态学特性存在着一定的差异,尽管三者均为湿地树种,但对湿地生境的适应性不同,毛赤杨为喜湿树种,而白桦和落叶松则为耐湿树种(牟长城等,2004),势必导致天然沼泽森林的年蓄积生长量沿过渡带环境梯度呈递减趋势;同时,由于排水使生境趋于中生化,有利于兴安落叶松的生长,使其在过渡带环境梯度的中上部生境地段具有较高的年蓄积生长量。
2.3 大兴安岭人工落叶松沼泽林植物多样性大兴安岭人工落叶松沼泽林与天然沼泽林的物种多样性沿过渡带环境梯度的变化趋势基本一致,呈现出逐渐增高的变化趋势(表 2)。在沼泽-森林过渡带下部生境上,物种丰富度和Shannon-Wiener多样性指数均较低,而在过渡带上部生境上两者均较高。但人工落叶松沼泽林与天然沼泽林的物种多样性存在较大的差异,并且与选用的多样性指数有密切关系。首先,人工落叶松沼泽林的物种丰富度指数普遍高于其相应生境地段的天然沼泽林。在过渡带0~100、100~200、200~300和300~400 m生境地段上,人工落叶松沼泽林的物种数量依次较天然沼泽林增加了9、25、14和12种,因此,人工落叶松沼泽林有利于多物种的共存。这主要是由于人工造林时排水整地使部分生境趋于中生化,导致中生植物大量侵入,并与原有湿生植物类群呈混杂状态而存在,势必导致群落物种丰富度的增加。其次,人工落叶松沼泽林的Shannon-Wiener多样性指数普遍低于其相应生境地段的天然沼泽林。在过渡带0~100 m生境地段上,人工落叶松沼泽林的群落Shannon-Wiener多样性指数较天然沼泽林群落下降了1.116 9;而在过渡带其他生境上,各群落的多样性指数平均值较天然沼泽林群落下降了0.576 2,其草本层下降了0.255 7,灌木层下降了1.02 2 0,乔木层下降了0.053 1。灌木层下降幅度最大,草本层下降幅度居中,而乔木层下降幅度最小。由此可见,灌木层和草本层多样性的减少是导致人工落叶松沼泽林Shannon-Wiene r多样性指数较天然沼泽林低的主要影响因素。再者,人工落叶松沼泽林的均匀度指数普遍低于其相应生境地段的天然沼泽林。人工落叶松沼泽林群落的Jsw均匀度指数,在过渡带0~100 m生境地段上,较天然沼泽群落下降了0.404 2;在过渡带其他生境上,各地段人工落叶松沼泽林群落的均匀度指数平均值较天然沼泽林群落下降了0.268 9,其草本层下降了0.284 7,灌木层下降了0.468 9,乔木层下降了0.075 2。也是灌木层下降幅度最大,草本层下降幅度居中,而乔木层下降幅度最小。这说明虽然人工落叶松沼泽林物种丰富度指数较高,但由于物种之间个体数量分布不均,从而导致群落的Shannon-Wiener多样性指数却较天然沼泽林有所下降。
|
|
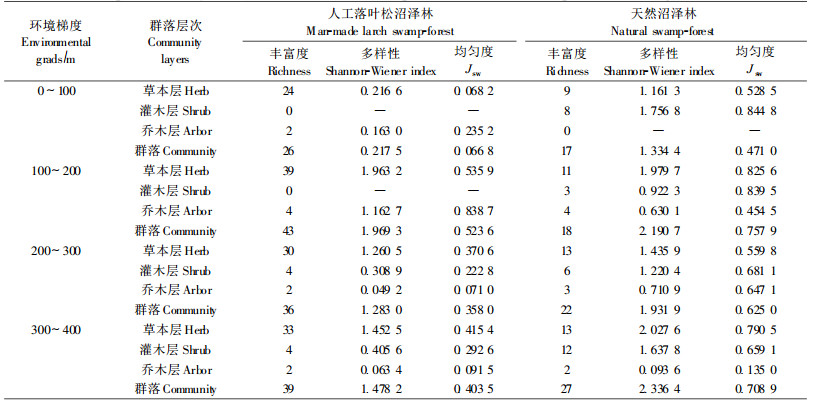
总体上看,在大兴安岭林区,排水造林改造的人工落叶松沼泽林与天然沼泽林相比较,尽管前者物种丰富度较高,但其物种多样性指数却与天然沼泽林存在较大的差距,在沼泽-森林过渡带各生境地段,其物种多样性指数较其相应生境地段天然沼泽林的下降幅度在0.221 4~0.858 2之间。这可能与人工林密度有关,由于人工林密度较大,林内透光性较差,不利于物种的侵入与生存,而在个别光照条件较好的局部地段上,却有较多的灌木与草本种类分布,这种分布上的不均匀性可能是导致人工落叶松沼泽林多样性指数降低的主要原因。因此,通过合理的林分密度管理与生境管理,创建一个林内透光条件良好,且积水与斑块镶嵌的生境格局,将有助于提高人工落叶松沼泽林的物种多样性。
2.4 大兴安岭人工落叶松沼泽林的环境变化人工排水造林改造后,人工落叶松沼泽林内环境与天然沼泽林差异较大,这种差异与环境梯度密切相关(表 3)。总体趋势为地下水位下降,下降幅度约为15.5~36.1 cm,泥炭层厚度减少,幅度约为9.6~25.2 cm,过渡带上部变化的程度较下部大。分析其原因,主要是由于排水整地抬升了沼泽地的微地势,改变了沼泽-森林过渡带的积水周期与积水量,改善了土壤的通气条件,增强了微生物的分解作用,加速了泥炭的分解过程,从而使泥炭层逐步转化为土壤层。此外,现有研究结果表明:沼泽地排水和整地对土壤的理化性质也具有较大的影响,土壤中速效氮、速效磷、速效钾都有不同程度的增加,排水时间越长,速效养分提高的幅度越大(白燕等,1996;张武文等,1996);且速效养分的增加促进了人工落叶松林林分的生长(李博生等,1996),因此,人工落叶松沼泽林的林分生产力较天然沼泽林有较大程度的提高。同时,地下水位与泥炭层的变化将会影响到沼泽-森林微生境的变化及其水文功能的发挥,从而进一步影响到树木的生长发育过程和植物多样性分布格局,具体影响机制尚有待进一步研究。
|
|

根据现有测算结果,对沼泽-森林过渡带进行人工造林改造,平均工程费用(包括抚育费)约为1 190元·hm-2(韩风海,2000);如果初植密度为3 000株·hm-2,按照2005年落叶松苗木(2年生)市场均价0.08元·株-1,则苗木费用为240元·hm-2,人工改造成本合计约为1 430元·hm-2,按5%的银行贷款利率计算,22年的成本为4 183元·hm-2。落叶松原木木材2005年的市场价(12~18 cm)为500元·m-3,依此价格,出材率按65%计算,人工改造的22年生落叶松沼泽林,除过渡带0~100 m生境地段的落叶松未达到中径材标准,无法依据常规计算外,在10 0~200、200~300、300~400 m的生境地段,可获得经济效益分别为10 138、31 784、4 0 335元·hm-2,投入产出比分别为1:2.4、1:7.6、1:9.6。由此可见,在沼泽-森林过渡带内培育落叶松人工林可获得较高经济效益,特别是在过渡带中上部生境地段可获得更高的经济效益。
3 结论与讨论1) 大兴安岭人工落叶松沼泽林与天然沼泽林在群落结构特征方面存在较大差异。天然沼泽林群落建群种沿沼泽-森林过渡带环境梯度发生明显的更替现象,依次分布着典型草本沼泽、毛赤杨沼泽林、白桦沼泽林和落叶松沼泽林,而人工落叶松沼泽林均以落叶松为优势种,且因灌木层和草本层发育不健全,不具备完整的垂直结构。这在很大程度上限制了人工落叶松沼泽林长期生产力维持、生物多样性保护及多种功能正常发挥,因此,有必要进一步探讨人工落叶松沼泽林群落合理结构及其形成机制。
2) 沼泽-森林过渡带环境梯度对其蓄积生长量具有重要影响,沿着沼泽到森林方向的环境梯度,人工落叶松沼泽林的蓄积生长量呈现逐渐增加的趋势,而天然沼泽森林的蓄积生长量却呈现逐渐减少的趋势。这一研究结论对于沼泽-森林利用和管理具有重要的指导意义,即应依据沼泽-森林过渡带环境梯度进行保护和利用,在过渡带0~100 m生境地段(典型沼泽生境地段)进行排水造林,林分蓄积生长量极低,经济效益极差,且导致植物多样性的下降及调蓄水功能的削弱,该区域不适宜发展林业;而在沼泽-森林过渡带其他生境地段(100 ~400 m生境地段),人工落叶松沼泽林的年蓄积生长量均较高(4.349 7~6.992 m3 ·hm-2a-1),可获得较高的经济效益,该区域是比较适宜发展林业的;但在沼泽-森林过渡带100 ~ 200 m生境地段,天然毛赤杨沼泽林年蓄积生长量为人工落叶松沼泽林的1.54倍,在该生境地段应以培育毛赤杨沼泽林为主。
3) 人工落叶松沼泽林与天然沼泽林的物种多样性存在较大差异。两者沿沼泽至森林方向过渡带环境梯度均呈现逐渐增高的变化趋势,但人工落叶松沼泽林物种丰富度指数较相应生境地段天然沼泽林群落高出9~25种,而其Shannon-Wiener指数和Jsw均匀度指数却较相应的天然沼泽林分别下降了0.221 4~0.858 2和0.234 3~0.404 2,因此,沼泽-森林排水造林尽管有利于多物种的共存,但由于各物种的个体数量分布不均匀导致了人工沼泽林的物种多样性较天然沼泽林有所降低。这可能与林分结构与林地微生境格局有密切关系,确切的影响机理尚有待进一步探讨。
4) 人工落叶松沼泽林与天然沼泽林的林地环境存在较大差异。其地下水位较天然沼泽林下降了15.5~36.1 cm,泥炭层厚度较天然沼泽林减少了9.6~25.2 cm,土壤中氮、磷、钾等速效养分含量增加,从而有利于落叶松的生长。但排水造林可能会影响沼泽-森林的水文功能及其生态系统内微生境的分布格局,从而影响到沼泽-森林的生态功能发挥及生物多样性的维持,这方面的影响机制及其影响效果也有待进一步系统研究。
5) 目前有关沼泽-森林的经营目标有3种选择:一是以发展高产农业和培育用材林为目标;二是作为自然保护区,保持原始状态;三是既能生产一部分木材,同时也要求保持它的结构和功能,既要能维持物种多样性,同时也要发挥它调节洪水和改善水质的水文功能。从我国实际效果来看,显然只有多目标的生态系统经营途径才是持久可行的(徐化成,2004)。从本项研究结果来看,沼泽-森林过渡带环境梯度是制约其生产力的主要因素,因此,多目标的经营应该是基于过渡带环境梯度的分类经营,在沼泽-森林过渡带的下部生境地段(典型沼泽生境地段)应保持原始状态,以发挥其水文调节及维持生物多样性功能;在沼泽-森林过渡带的中下部生境地段,应以培育毛赤杨沼泽林为主;在沼泽-森林过渡带的中上部生境地段则可以用来培育人工落叶松沼泽林。这样既能保证获得较高的林分生产力,又能维持沼泽-森林群落的生物多样性及其正常的水文功能。
白燕, 郎惠卿. 1996. 森林沼泽改造后泥炭土的速效养分变化. 东北师范大学学报:自然科学版, 28(4): 105-110. |
韩风海. 2000. 低湿地改造与天然更新. 林业科技通讯, (8): 43-44. DOI:10.3969/j.issn.1671-4938.2000.08.021 |
郎惠卿. 1996. 中国湿地与保护//国家林业局保护司. 中国湿地保护与持续利用研究论文集. 北京: 中国林业出版社, 63-67
|
朗惠卿, 赵魁义, 陈克林, 等. 1999. 中国湿地植被. 北京: 科学出版社, 1-73.
|
李博生, 丁国栋, 张武文, 等. 1996. 大兴安岭水湿地兴安落叶松蒸腾作用及其应用的研究. 内蒙古林学院学报:自然科学版, 18(4): 18-23. |
李坚. 1994. 现有林经营管理导论. 哈尔滨: 东北林业大学出版社, 41-60.
|
马学慧, 牛焕光. 1991. 中国的沼泽. 北京: 科学出版社, 73-88.
|
牟长城, 韩士杰, 罗菊春, 等. 2001a. 长白山森林/沼泽生态交错带群落和环境梯度分析. 应用生态学报, 12(1): 1-7. |
牟长城, 韩士杰, 罗菊春, 等. 2001b. 长白山森林沼泽生态交错带群落演替规律研究(Ⅰ):白桦沼泽群落演替规律研究. 北京林业大学学报, 23(1): 1-6. |
牟长城, 罗菊春, 王襄平, 等. 1998. 长白山林区森林沼泽交错群落的植物多样性. 生物多样性, 6(2): 132-137. DOI:10.3321/j.issn:1005-0094.1998.02.008 |
牟长城, 万书成, 苏平, 等. 2004. 长白山毛赤杨和白桦沼泽生态交错带群落生物量分布格局. 应用生态学报, 15(12): 2211-2216. DOI:10.3321/j.issn:1001-9332.2004.12.003 |
牟长城. 2003. 长白山落叶松和白桦-沼泽生态交错带群落演替规律研究. 应用生态学报, 14(11): 1813-1819. DOI:10.3321/j.issn:1001-9332.2003.11.001 |
王荣芬. 1994. 我国沼泽资源开发利用现状. 地理科学, 14(3): 250-259. |
徐化成. 2004. 森林生态与生态系统经营. 北京: 化学工业出版社, 396-397.
|
许红梅, 许林书. 2000. 小兴安岭泥炭沼泽土排水改造前后营养元素生物循环的实验研究. 东北师范大学学报:自然科学版, 32(2): 73-77. |
张武文, 马秀珍, 丁国栋. 1995a. 大兴安岭水湿地人工幼林生长规律的初步探讨. 内蒙古林业科技, (1): 5-9. |
张武文, 邹庆普, 周文恒. 1995b. 大兴安岭水湿地类型划分及改造利用. 内蒙古林学院学报:自然科学版, 17(1): 23-29. |
张武文, 马秀珍, 丁国栋. 1996. 大兴安岭水湿地改造对土壤和植被的影响. 东北林业大学学报, 24(6): 9-13. |
Barnes W J. 1985. Population dynamics of woody plants on a river island. Canadian Journal of Botany, 63: 647-655. DOI:10.1139/b85-081 |
Hodges J D. 1995. The southern bottomland hardwood region and brown loam bluffs subregion//Barrett J W. Regional Silviculture of the United States (3rd). New York: Wiley and Sons, 227-269
|
Hodges J D. 1997. Development and ecology of bottomland hardwood sites. Forest Ecology and management, 90: 117-125. DOI:10.1016/S0378-1127(96)03906-0 |
Hodges J D. 1998. Minor alluvial floodplains//Messina M G, Conner W H. Southern forested wetlands: ecology and management. Boca Raton, Florida: Lewis Publishers, 325-341
|
Kellison R C, Young M J, Brahan R R, et al. 1998. Major alluvial floodplains//Messina M G, Conner W. Southern Forested Wetlands: Ecology and Management. Boca Raton, Florida: Lewis Publishers, 291-324
|
MacDonald P O, Frayer W E, Clauser J K. 1979. Documentation, chronology, and future projections of bottomland hardwood habitat losses in the Lower Mississippi Alluvial Plain. Volum 1. US Fish and Wildlife Service, Washington
|
Ohlson M, Soderstrom L, Hornberg G, et al. 1997. Habitat qualities versus long-term continuity as determinants of biodiversity boreal old-growth swamp forests. Biological Conservation, 81: 221-231. DOI:10.1016/S0006-3207(97)00001-3 |
Schnitzler A. 1994. European alluvial hardwood forests of large floodplains. Journal of Biogeography, 21: 604-623. |
Schnitzler A. 1995a. Community ecology of arboreal lianas in gallery forests of the Rhine valley, France. Acta Oecologia, 16: 219-236. |
Schnitzler A. 1995b. Successional status of trees in gallery forest along the river Rhine. Journal of Vegetation Science, 6: 479-486. DOI:10.2307/3236346 |
Schnitzler A, Hale B W, Alsum E. 2005. Biodiversity of floodplain forests in Europe and eastern North America: a comparative study of the Rhine and Mississippi Valleys. Biodiversity and Conservation, 14: 97-117. DOI:10.1007/s10531-005-4056-2 |
Sharitz R R, Mitsch W J. 1993. Southern Floodplain Forests//Martin W H, Boyce S G, Echternacht A C. Biodiversity of the Southeastern United States: Lowland Terrestrial Communities. New York: John Wiley and Sons, 311-372
|
Shepard J P. 1995. Opportunities: reforesting marginal agricultural land. Forest Farmer, 54: 7-9. |
Stanturf J A, Schweitzer C J, Gardiner E S. 1998. Afforestation of marginal agricultural land in the Lower Mississippi River Alluvial Valley. USA. Silva Fennica, 32: 281-297. |
Stanturf J A, Schoenholtz S H, Schweitzer G J, et al. 2001. Achieving restoration success: Myths in bottomland hardwood forests. Restoration Ecology, 9(2): 189-200. DOI:10.1046/j.1526-100x.2001.009002189.x |