 2007, Vol. 43
2007, Vol. 43文章信息
- 柳新红, 何小勇, 苏冬梅, 魏来, 袁德义, 王军峰.
- Liu Xinhong, He Xiaoyong, Su Dongmei, Wei Lai, Yuan Deyi, Wang Junfeng.
- 翅荚木种源抗寒性综合评价体系的构建与应用
- Establishment and Application of Integrated Assessment System on Cold Resistance of Different Zenia insignis Provenances
- 林业科学, 2007, 43(10): 45-50.
- Scientia Silvae Sinicae, 2007, 43(10): 45-50.
-
文章历史
- 收稿日期:2007-07-31
-
作者相关文章


2. 丽水市科学技术协会 丽水 323000;
3. 中南林业科技大学 株洲 412006;
4. 丽水市林业科学研究所 丽水 323000
2. Lishui Science and Technology Association of Zhejiang Province Lishui 323000;
3. Central South University of Forestry and Technology Zhuzhou 412006;
4. Lishui Forestry Institute of Zhejiang Province Lishui 323000
翅荚木( Zenia insignis)又名任豆树、任木、砍头树,属苏木科(Caesalpiniaceae)翅荚木属(Zenia)落叶高大乔木,自然分布于桂、粤、湘、黔、滇等省(区)(陈永密等,1989),是国家Ⅱ级保护野生植物(傅立国,1992),世界自然保护联盟(IUCN)红色名录中的近危种。翅荚木树干通直、生长迅速、萌芽性强、天然更新好、抗性强、耐瘠薄,20世纪80年代先后被湖南、福建、江西、浙江、河北、四川等地引种,表现出比当地乡土树种更好的速生性(覃志刚等,1992;何义发等,1996;童方平等,2005)。翅荚木是热带亚洲分布的植物(吴征镒,1991),引种后冬季遇低温时易受冻害,苗期和幼林期尤为严重,但不同种源之间的冻害差异极大(陈亮明等,1997)。因此,选育翅荚木抗寒良种并准确鉴定良种的抗寒性尤为重要,而目前相关研究尚未展开。
林木的抗寒性综合评估是现代林木抗性育种工作的一项重要内容。关于林木抗寒性的评价,国内外学者已经提出了一大批与抗寒性测定有关的形态指标和生理生化指标(张钢,2005;徐燕等,2007)。由于林木的抗寒性是许多数量或质量遗传基因综合作用累加的结果,每一个与抗寒性有关的性状对林木的抗寒性都起一定的作用,但这种作用是微效的。所以,单一指标评价翅荚木抗寒性具有片面性,用多个指标综合评价翅荚木的抗寒性才较为可靠。本文在对翅荚木的生理生化物质、叶片解剖、田间试验、恢复生长等进行系统研究的基础上,利用主成分分析的方法,探讨适宜的评价指标,建立综合评价体系,对不同翅荚木种源进行抗寒性鉴定,以期为翅荚木抗寒良种选育提供理论依据。
1 材料与方法 1.1 供试材料翅荚木6个种源,分别于2004年采集于广西壮族自治区的靖西和忻城、广东省的英德、湖南省的通道和江华、贵州省的兴义。种子采集后自然干燥,密封贮藏。2005—2006年每年4月中旬播种,小苗移植于中南林业科技大学湖南省株洲市教学基地和浙江省丽水市林业科学研究所苗木基地,其中种植时前者各种源随机排列,后者按随机区组设计,重复3次。苗木生长期间施肥2次(用尿素150 kg·hm-2),锄草2次,防治蚜虫1次。
1.2 指标测定 1.2.1 抗寒生理指标的测定生理指标测定材料为1年生苗木,2007年1月中旬在中南林业科技大学湖南省株洲市教学基地采样。每种源选取生长一致的苗木35株,每株截取60~80 cm的健壮枝条,洗净擦干。每种源分7组分别用湿纱布将枝条包裹后装入塑料袋中,置于低温冰箱内进行梯度降温冷冻处理。梯度降温设4、0、-4、-8、-12、-16、-20 ℃共7个处理,以4 ℃为对照(CK)。降温速度为1 ℃·h-1。降到所需温度维持24 h后取出1组枝条升温到4 ℃,其余枝条继续进行降温处理。各处理后的枝条取上部2~3节避开芽眼剪成0.2~0.3 cm的小段,混合均匀后用于各项生理指标测定(重复3次),其余部分用于恢复性生长试验。其中相对电导率(relative electric condu ctivity,REC)采用EC200型电导/TDS/盐度测定仪测定(王晶英等,2003),并推算半致死温度(semi-lethal temperature,LT50)(史清华等,2003);超氧化物歧化酶SOD酶活性用氮蓝四唑(NBT)的光化还原法测定(中国科学院上海植物生理研究所,1999);可溶性蛋白含量用考马斯亮蓝G-250染色法测定(王晶英等,2003);可溶性糖含量用蒽酮法测定(王晶英等,2003);丙二醛(MDA)含量用硫代巴比妥酸(TBA)显色法测定(汤章诚,1999);恢复生长时将低温处理后的枝条剪成15 cm长的枝段进行水插,放入光照培养箱中培养,温度(25±2) ℃,光照14 h/黑暗10 h,30 d后统计萌芽展叶情况,分种源统计萌芽率,计算相对萌芽率,即各处理的芽萌发率与对照(CK)的芽萌发率的比例(缴丽莉等,2006)。
1.2.2 叶片解剖采样与制片分析2005年8月上旬从中南林业科技大学湖南省株洲市教学基地的1年生翅荚木苗木上采集叶片样本。每种源随机选取5株,每株统一选取离地高50 cm处的发育成熟的羽叶,摘取中部的小叶1片作样本。每叶从小叶的中部切取1 cm ×0.5 cm制样,用FAA固定液固定,常规石蜡制片法切片;切片厚度12 μm,番红、固绿双重染色,中性胶固封;制片后,每叶取一张代表性切片在Motic DMB5型数码生物显微镜下观察照相,用数码测距软件Motic Images Advanced 3.0测量叶片表皮层、栅栏组织和海绵组织的厚度,每项指标重复测定10次,取平均值计算细胞结构紧密度(cell tense ratio,CTR)和疏松度(spongy ratio,SR),其中:CTR=(栅栏组织厚度+下部紧密组织厚度)/叶片厚度×100%;SR =海绵组织厚度/叶片厚度×100 %。每叶从小叶中脉一侧切取1 cm×0.5 cm制样,放入10 %硝酸+10 %三氧化铬混合溶液(40~50 ℃)充分氧化,离析表皮,修片后0.5%番红水溶液染色,二甲苯脱水,中性胶固封;每片选取10个视野显微观测气孔大小和密度,测量表皮毛长度和数量,计算每叶的平均值(刘世彪等,2004)。
1.2.3 生长性状指标的测定生长性状指标测定在浙江省丽水市林业科学研究所苗木基地进行。种子收齐后,每种源采用以十字对角取样法分别抽取4×100粒测定千粒质量,随机取30粒种子测定其宽和高。2006年9月,在丽水当年播种苗中每小区随机选取10株,每株选取中部羽叶测定长度和小叶数,并选取中部1小叶扫描仪扫入,利用图像处理软件Photoshop按像素测定小叶长度、宽度和面积(左欣等,2006)。2007年2月,每小区随机挖取3株生长正常的苗木,分别测量苗高、地径、根鲜质量、茎鲜质量等指标。
1.3 数据处理主成分分析、聚类分析等相关分析采用统计软件SPSS13.0进行。
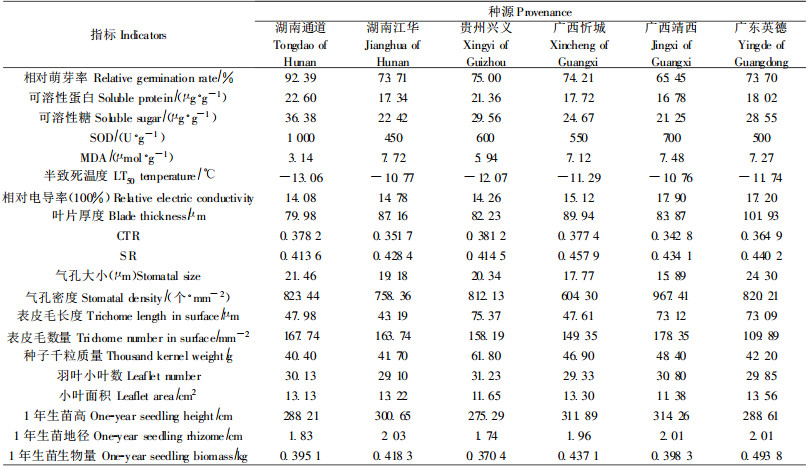
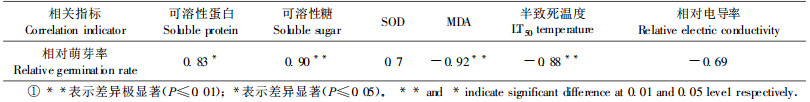
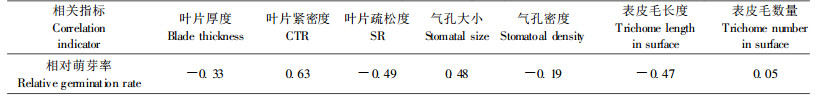
2 结果与分析 2.1 抗寒指标相关性分析环境胁迫对植物体的影响是多方面的,但最终体现在植物的生长和发育上(徐文铎等,1993)。芽是植物对低温最敏感的部位,经过冷冻处理后,各种源枝条均受到不同程度的伤害。随着处理温度的降低,枝条萌芽率逐渐降低。经- 8 ℃低温处理后,大部分种源的萌芽率明显下降,且种源间差异较大;经-12 ℃低温处理后的各个种源枝条均未萌发,说明-12 ℃将对翅荚木苗木造生严重冻害,直至死亡。因此,研究以-8 ℃低温处理后的各种源枝条相对萌芽率作为翅荚木抗寒性的衡量指标。应用相关分析法,计算各种源经-8 ℃低温处理后的生理指标(可溶性蛋白、可溶性糖、SOD、MDA含量和相对电导率)、半致死温度、叶片解剖性状指标(叶片厚度、CTR、SR、气孔大小和密度、表皮毛长度和数量)、生长形态指标(千粒质量、小叶数、小叶面积、苗高、地径、生物量)(表 1)与相对萌芽率的相关性,结果如表 2~4。
|
|
|
|
|
|
|
|

从表 2~4可以看出:翅荚木种源的可溶性糖含量、MDA含量、枝条的半致死温度与抗寒性极显著相关,可溶性蛋白含量与抗寒性显著相关。叶片解剖结构指标中与抗寒性相关最大的是叶片紧密度,生长形态指标与抗寒性相关最大的是生物量。
2.2 抗寒综合评价体系的建立 2.2.1 抗寒指标的选取根据相关性分析的结果,取极显著或显著相关的可溶性糖含量、MDA含量、枝条的半致死温度、可溶性蛋白含量作为生理生化指标,相关性最好的CTR值作为叶片解剖结构指标,生物量作为生长形态指标建立翅荚木种源抗寒性的评价体系,这些指标可以在一定程度从生理、组织结构、形态上反映植物的抗寒性。
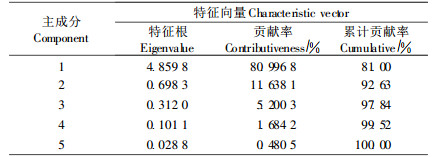
2.2.2 抗寒指标权重的确定由于单一指标评价植物抗寒性存在片面性,多个指标进行综合评价才较为可靠。但不同指标之间的关系复杂,并且不同指标的重要性也不相同,因此需要进行主成分分析,结果如表 5。从表 5可以看出,1~5主成分提取特征向量贡献率分别为80.996 8%~0. 480 5 %,累计贡献率达到100.00%。一般认为如果主成分分析中所提取主成分的特征值能达到70%以上的贡献率,就可以用这个或几个主成分对事物的属性进行概括性分析,基本可以得出影响事物性质的主要因素。本研究第一主成分的贡献率达到了81.00%,可以代表原始指标的绝大部分信息。因此,以第一主成分分析各指标的负荷量,计算各指标对翅荚木抗寒性的作用大小,确定权重Wi,结果见表 6。从表 6可以看出,对翅荚木抗寒性影响最大的指标是根据电解质透出率达50%得出的半致死温度,以下依次是可溶性蛋白含量、可溶性糖含量、MDA含量、叶片结构紧密度和生物量。
|
|
|
|
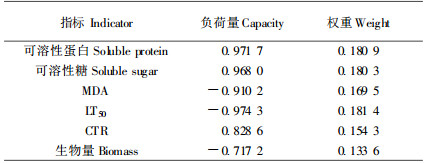
由于各指标的性质、单位和数量不同,因此,需要对这些指标进行标准数量化。各指标变化具有连续性质,故采用连续性质的隶属度函数,并根据主成分因子负荷量的正负性,确定隶属度函数的升降性。MDA、LT 50、生物量采用降型分布函数:f(xi)=(xi max-xij)/(xi max-xi min);可溶性蛋白、可溶性糖和CTR采用升型分布函数:f(xi)=(xij-xi min)/(xi max-xi min),其中f(xi)表示各指标的隶属度值,xij表示各指标值,xi max和xi min分别表示第i项指标的最大值和最小值。各种源相关指标的隶属度值见表 7。
|
|
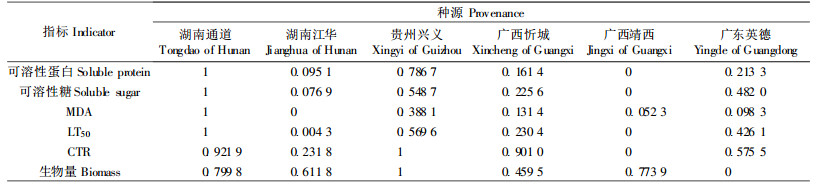
根据加乘法则:I=Wi×f(xi) (其中:Wi为抗寒性指标的权重向量;f(xi)为各指标的隶属度值),对各指标进行合成,计算各种源的抗寒性综合指数I。根据综合指数的大小,可知其抗寒性强弱的排序为:湖南通道>贵州兴义>广西忻城>广东英德>湖南江华>广西靖西(表 8)。
|
|

把各种源的抗寒综合指数值用类平均法进行聚类分析,可将其划分为3类:湖南通道和贵州兴义种源为一类,抗寒性较好;广西忻城和广东英德种源为一类,抗寒性一般;湖南江华和广西靖西种源为一类,抗寒性较差(图 1)。

|
图 1 不同抗寒性的翅荚木种源聚类图 Fig. 1 Cluster of Z. insignis provenance with different cold resistance capacity 1. 广西靖西 Jingxi of Guangxi;2. 湖南江华 Jianghua of Hunan; 3. 广东英德 Yingde of Guangdong;4. 广西忻城 Xincheng of Guangxi; 5. 贵州兴义 Xingyi of Guizhou;6. 湖南通道 Tongdao of Hunan. |
植物受低温胁迫后的恢复生长情况直接反映了植物的抗寒性,本文根据与恢复生长相关性大小确定了6个翅荚木抗寒指标。其中,可溶性蛋白质具有强亲水性,其含量的增加可显著加强细胞的保水力,提高细胞内的束缚水含量和原生质弹性,降低冰点,防止细胞内结冰,提高植物组织的抗寒性;而低温下可溶性糖含量常常与可溶性蛋白含量密切相关,它们常作为鉴定植物抗寒性的指标;MDA是膜脂过氧化作用的最终产物,也是衡量膜系统受害的重要指标之一;电导率的变化反映出质膜伤害程度和所测材料的抗寒性的大小,电解质透出率达50 %得出的半致死温度能够更加准确的反映植物的抗寒性;植物叶片的CTR值与抗寒性密切相关已在许多植物上已经得到证明。它们可以一定程度地从生理、组织结构、形态上反映植物抗寒性的大小。
由于植物体是一个统一的有机体,如果使用性质相同或相互关联的指标来综合评价其抗寒性可能会使结果造成偏差。本文通过相关性分析确定衡量翅荚木单项抗寒能力大小的指标,用主成分分析法将原来个数较多且彼此关系错综复杂的指标转换成综合的主成分,并根据主成分的各指标的负荷量,计算各指标对抗寒性作用的大小,确定权重,再运用隶属函数法进行综合评价,使结果更加可靠。根据综合指数的大小,可知翅荚木不同种源的抗寒性强弱的排序为:湖南通道>贵州兴义>广西忻城>广东英德>湖南江华>广西靖西,这与我们引种育苗时冻害发生情况基本一致。2005年12月,浙江省丽水市连续七天的日最低气温均低于零度,最低气温达-6.1 ℃,苗木平均冻害率达到了71.88 %,而湖南通道种源的冻害率仅为4.83 %,极显著低于其他种源,说明了湖南通道种源比其它种源具有较好的抗寒性。从地理分布来看,湖南通道纬度较高,贵州兴义海拔较高,这两地的气温较低,其种源抗寒性较好,这可能是林木适应自然选择的结果。
影响植物抗寒性的因素较多,其抗寒机理错综复杂。目前,国外测定木本植物抗寒性有全株冰冻测试法(Tanaka et al., 1997)、电解质渗出率法(Deans et al., 1995)、叶绿素荧光法(Maxwell et al., 2000)、热分析法(Kang et al., 1998)等方法,国内在形态结构、膜脂的成分及蛋白质、抗寒生理生化、抗寒基因等方面的研究已经取得显著进展(马英姿等,2005)。具体评定翅荚木种源抗寒性时,还应多方面综合考虑,同时结合田间观察,才能为全面评价其抗寒能力提供可靠的依据。
陈亮明, 陈永密, 张巧琴. 1997. 翅荚木引种栽培耐寒力的调查研究. 林业科技开发, (3): 37-38. |
陈永密, 张巧琴, 张石玉, 等. 1989. 珍稀树种——翅荚木. 福州: 福建教育出版社.
|
傅立国. 1992. 中国植物红皮书——稀有濒危植物. 第一册. 北京: 科学出版社.
|
何义发, 严昌荣. 1996. 珍稀树种——翅荚木在恩施山地引种试验初报. 湖北林业科技, (1): 24-26. |
缴丽莉, 路丙社, 白志英, 等. 2006. 四种园林树木抗寒性的比较分析. 园艺学报, 33(3): 667-670. DOI:10.3321/j.issn:0513-353X.2006.03.046 |
刘世彪, 陈青, 胡正海. 2004. 7种番荔枝果树的叶片结构及其与抗寒性关系研究. 果树科学, 21(3): 241-246. DOI:10.3969/j.issn.1009-9980.2004.03.013 |
马英姿, 梁文斌, 陈建华. 2005. 经济植物的抗寒性研究进展. 经济林研究, 23(4): 89-94. DOI:10.3969/j.issn.1003-8981.2005.04.025 |
史清华, 高建社, 王军, 等. 2003. 5个杨树无性系抗寒性的测定与评价. 西北植物学报, 23(11): 1937-1941. DOI:10.3321/j.issn:1000-4025.2003.11.015 |
覃志刚, 邱进贤. 1992. 翅荚木引种栽培的研究. 四川林业科技, 13(1): 59-63. |
汤章诚. 1999. 现代植物生理学实验指导. 北京: 科学出版社, 305-306.
|
童方平, 吴际友, 龙应忠, 等. 2005. 珍稀速生树种翅荚木栽培技术研究. 湖南林业科技, 32(4): 13-15. DOI:10.3969/j.issn.1003-5710.2005.04.004 |
王晶英, 敖红, 张杰, 等. 2003. 植物生理生化实验技术与原理. 哈尔滨: 东北林业大学出版社.
|
吴征镒. 1991. 中国种子植物属的分布区类型. 云南植物研究, (增刊Ⅳ): 1-319. |
徐文铎, 郑元润, 刘广田. 1993. 内蒙古沙地云杉生长与生态条件关系的研究. 应用生态学报, 4(4): 368-373. DOI:10.3321/j.issn:1001-9332.1993.04.016 |
徐燕, 薛立, 屈明. 2007. 植物抗寒性的生理生态学机制研究进展. 林业科学, 43(4): 88-94. |
张钢. 2005. 国外木本植物抗寒性测定方法综述. 世界林业研究, 18(5): 14-20. DOI:10.3969/j.issn.1001-4241.2005.05.004 |
中国科学院上海植物生理研究所. 1999. 现代植物生理学实验指南. 北京: 科学出版社, 314-315.
|
左欣, 韩斌, 程嘉林. 2006. 基于数字图像处理的植物叶面积测量方法. 计算机工程与应用, 42(27): 194-196. DOI:10.3321/j.issn:1002-8331.2006.27.059 |
Deans J D, Billington F J, Harvey F J, et al. 1995. Assessment of frost damage to leafless stem tissues of Quercus petraea: a reappraisal of the method of relative conductivity. Forestry, 68: 25-34. DOI:10.1093/forestry/68.1.25 |
Kang S K, Motosugi H, Yonemori K, et al. 1998. Supercooling characterics of some deciduous fruit trees as related to water movement within the bud. Journal of Horticultural Science and Biotechnology, 73(2): 165-172. DOI:10.1080/14620316.1998.11510960 |
Maxwell K, Johnson G N. 2000. Chlorophyll fluorescence-a practical guide. Journal of Experimental Botany, 51: 659-668. DOI:10.1093/jexbot/51.345.659 |
Tanaka Y, Brotherton P, Hostetter S, et al. 1997. The operational planting stock quality testing program at Weyerhaeuser. New Forests, 13: 423-437. DOI:10.1023/A:1006567231885 |