 2007, Vol. 43
2007, Vol. 43文章信息
- 刘兆刚, 李凤日.
- Liu Zhaogang, Li Fengri.
- 樟子松人工林树冠内一级枝条空间的分布规律
- Modeling of Spatial Distribution of Primary Branches Within the Crowns of Pinus sylvestris Stands
- 林业科学, 2007, 43(10): 19-27.
- Scientia Silvae Sinicae, 2007, 43(10): 19-27.
-
文章历史
- 收稿日期:2006-12-08
-
作者相关文章



树木的地上部分主要由树干、树枝、树叶等部分组成,其中枝、叶组成的树冠是高大乔木进行光合作用、呼吸作用、蒸腾作用等一系列生理活动的主要部位,其大小、结构、形状及其在林分中的分布形式直接决定了树木个体的各组分生物量、生长活力和生产力,并反映了林木在林分中的长势情况。组成树冠枝条的主要功能有2个:一是作为叶片的支撑体,决定着叶片的空间分布;二是连接叶片与树木整体,担负着它们之间的水分和营养物质的运输。枝条和叶片形成的空间排布格局作为树冠的构成单位,对它的清楚认识是了解树木生长发育的前提(马克明等,2000)。
一级侧枝是构成树的骨架,它的长短及其空间排列对树的形状起着支配作用,因此枝条的生长规律是研究树冠的结构和功能以及树木有机物生产量的基础。国外许多学者在树冠形状和结构的研究上都应用了枝条等树冠变量。猪濑光雄(1982)利用枝解析的方法分析过冷杉(Abies fabri)侧枝的径向生长和材积生长。铃木悌司等(1992)研究了北海道的代表针叶树库页冷杉(Abies sachalinensis)的枝生长模型(一次枝、二次枝以下的枝生长模型),得出轮生枝个数是以5为平均值、标准偏差1.02的正态分布,从枝的着枝角度、枝生长量、枯枝着枝年份等方面进行了比较细致地描述。有学者认为树干顶端新枝的数量受林地密度的影响(Carter et al., 1986),而也有研究表明每个轮枝内的枝条数量主要与树干的年高生长有关(Collin et al., 1992)。Hashimoto(1990; 1991)用枝条的属性因子研究了树冠形态的变化,认为光环境是影响树冠形状和结构的主要因素。Maguire等(1991)应用林分变量和相对着枝深度建立了树冠内主要枝条基径的预估模型,Tombleson等(1990)研究了林分变量如立地和立木度对树木枝条大小的影响,但是这些只研究最大的枝条或最大枝条的平均值,而所有枝条的大小和位置依然研究很少。Maguire等(1990)为花旗松(Pseudotsuga menziesii)幼龄林建立了各轮枝枝条数量的预估模型,并分析了各轮枝内枝条的相对频数分布,构造了模型来预测花旗松每年一级新生枝条基径和垂直分布,但枝条的方位角分布没有研究;Doruska等(1994)所建模型包括火炬松(Pinus taeda)人工林总枝条数、各轮枝枝条数及其水平分布模型,但他们并没有建立有关树冠内枝条垂直分布的模型。
近些年来,国内也有许多学者对树冠结构进行了专项研究。韩兴吉(1985)探讨了油松(Pinus tabulaeformis)树冠枝生长规律。赵天傍(1990)对望春玉兰(Magnolia biondii)幼树的枝条分布状态、侧枝数量、着枝角大小和轮枝间距离等一般结构特点进行了分析研究。祝宁等(1995)对药用植物刺五加(Acanthopanax senticosus)的构件和基本结构进行了统计分析。臧润国等(1998)初步分析了热带树木的基本结构和热带树木结构的总体特点和基本规律。李俊清等(2001)研究了欧洲水青冈(Fagus sylvatica)的结构与形态多样性关系。李凤日等(1996)、李凤日(2004)、Li等(2004)研究了长白落叶松(Larix olgensis)的树冠形状、结构及林分动态。刘兆刚等(2005)对樟子松(Pinus sylvestris)人工林树冠结构的分形进行了分析。
研究树冠结构规律特征对于深入研究树木的生长及其进行树冠的动态模拟具有重要意义。要描述樟子松的树冠结构,必须对组成树冠各结构因子及其之间的关系进行深入研究,通过定量和定性分析建立它们之间的数学模型。由于树冠变量的直接测定存在着极大的困难,因此本文的研究目的就是应用枝解析技术,利用数学模型和统计分析方法建立描述樟子松的树冠一级枝条数目预估模型,并探讨树冠枝条空间分布规律,为进一步研究樟子松人工林树冠结构规律及动态生长模拟提供理论基础。
1 材料与方法 1.1 试验材料分别于2002和2003年在东北林业大学帽儿山实验林场选择不同年龄、不同立地和不同密度的樟子松人工林中有代表性的林分,设置固定标准地15块(其中2002年设置7块,2003年设置8块),所选林分均未经过间伐,生长正常。在每块标准地选取冠形良好、无病虫害的解析木3~5株。
将解析样木仔细伐倒以免损坏树冠,以至少有1个活枝的轮枝的位置来确定树冠的基部。将解析木的树干严格按1 m区分段进行区分,并在每个区分段的中央位置锯取树干解析圆盘。树冠部分也严格按1 m区分段进行区分,树梢在一般情况下是不足1 m的梢头。从树梢端开始,在每一树冠区分段内分别轮枝逐枝进行编号,对每轮枝内的全部枝条进行枝条因子测定,包括:总着枝深度(DINC)、枝条的方位角(Φ)、着枝角度(θ)、基径(BD)、枝长(BL)和弦长(BCL)、弓高(BAH)等。
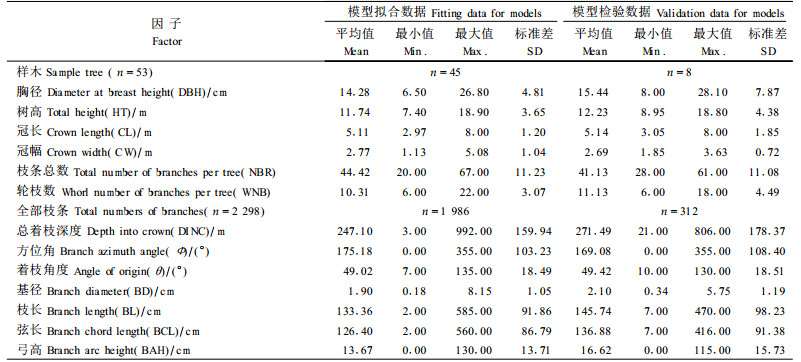
53株样木总共测量2 298个活枝条,其中作为拟合数据的枝条有1 986个,检验数据的枝条312个(表 1)。
|
|
应用45株拟合解析木分别统计了每株樟子松树冠内的所有活枝数(Nb),并将年龄(AGE)、胸径(DBH)、树高(HT)、冠长(CL)、冠长率(CL%)、枝下高(HCB)及林分密度(SD)、地位指数(SI)、郁闭度等林木和林分变量及变量的形式变换作为自变量来拟合各种模型,利用比较拟合优度,确定最佳模型,建立樟子松人工林树冠内一级枝条活枝总个数的预估模型。
1.2.2 枝条的空间分布1) 枝条的垂直分布 探讨枝条的垂直分布可以从2个方面进行研究:一是预估轮枝的高度模型;二是确定每轮内枝条的数目。
由于樟子松树木生长的特点,每年只形成一轮枝条,且每轮枝条在树干上的高度没有明显差别,因此预估轮枝的高度与预估该轮枝内单个枝条的高度是一致的。确定轮枝的高度尝试采用下列2种方法:一是通过建立轮枝高度与轮枝序、林木变量和林分变量的回归模型;另一种方法是将相对着枝深度(RDINC)划分为10个等级,探讨每个等级内枝条频数分布情况;然后利用Weibull分布、Gamma分布、正态分布等进行拟合,得出最优的分布函数,并探讨分布函数的估计参数与林分和树木变量的关系。
确定每轮枝条内的数目主要有2种途径:一是建立轮枝内枝条数目与林木、林分及枝条变量的模型;二是描述性分析轮枝内枝条数量的分布情况。
2) 枝条的水平分布 研究枝条的水平分布主要包括2个方面的内容:一是探讨枝条的方位角的分布;二是研究具有相同枝条数相邻轮枝间枝条方位角的关系。
由于树木的每轮枝条在树干上的位置是呈环形分布的,因此由Batschelet(1981)和Mardia(1972)提供的圆形分布统计量是用来分析这类数据有用的工具。均匀圆形分布可用下列函数表示
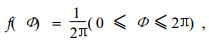
|
式中Φ为弧度。
① 圆形分布的平均角及其假设检验
设有n个角度α1, α2, …, αn,令a表示角的样本均数,简称平均角,其计算公式为
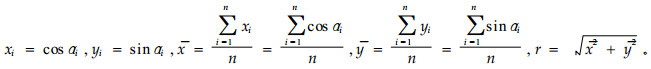
|
(1) |

|
(2) |
角的标准差公式为

|
(3) |
当一组数据中所有αi都等于同一数值,则这组数据无变异,S=0,而r=1;当一组数据中的αi均匀地分布在圆周上,则r=0,而S则因为平均角不存在而无法计算;但当r趋向于0时,S趋向于无穷大。r的取值的范围在0~1之间,S值的范围在0~∞之间,S可称为圆标准差或平均角离差,简称角离差,通常以r表示角度资料的集中趋势量度,S表示其离散趋势量度。
当基本参数计算处理后,就可进一步进行平均角均匀性检验。检验是基于所有αi都是均匀分布在圆周的一个总体,其集中趋势量度ρ=0,但在此总体中随机抽取一个样本,所得ρ的估计值不一定为0,因此,当同一个样本资料算得平均角a和r后,此a是否有意义(即是否来自ρ=0的总体)必须进行均匀性检验。
此时,H0:ρ=0即为均匀分布,不存在平均角;H1:ρ≠0即为非均匀分布,存在平均角。
均匀性检验方法是:根据样本大小n算得的r查表,如r大于或等于表中界值,则p≤α, 即在相应的水平上拒绝H0,表示存在集中趋势,平均角有意义; 如r小于表中界值,则p>α则拒绝H0,认为是均匀分布,不存在集中趋势,故平均角无意义。
应用53株树中的每株树分别确定枝条方位角是否符合均匀圆形分布,均值向量是否存在集中趋势度量,枝条平均方位角是否意义。
② 圆-圆相关性检验
当观察到n对角度数据(αi, βi)时,可以研究α和β之间的相关性,称为圆-圆相关性。其计算方法与圆形统计量是否均匀分布有关,当α和β都呈均匀分布时,可用H检验法来检验其相关性。其相关系数的计算公式为

|
(4) |
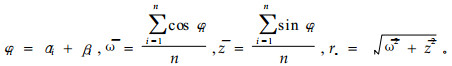
|
(5) |
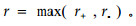
|
(6) |
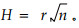
|
(7) |
圆-圆相关分析要求成对样本角度数据符合均匀圆形分布。本文选择具有相邻的轮枝,且每轮枝具有相同数目的枝条。正相关和负相关同时检验用来确定相关系数,正相关表明当成对的角度一个增加时,另一个也增加;负相关正好相反。每一成对轮枝的枝条(相邻的具有相同数目的枝条)按方位角排序后,按上式计算匹配枝条的角度差(和),然后计算相关系数。
首先分别样木统计具有相同枝条数目的相邻轮枝的匹配对数,然后应用DPS7.05统计软件的圆-圆相关分析确定相邻轮枝之间的相关系数。
2 结果与分析 2.1 一级枝条个数预估模型通过每株树木实测总枝条个数与林木及林分变量的简单相关分析发现,樟子松人工林的一级枝条总个数(Nb)只与冠长(CL)有关。试图引进其他的林木和林分因子,如胸径、树高、年龄等,但方程均没有明显地改进,因为这些变量提供的信息已经包含在树冠冠长变量中。通过模型的优选,本文应用冠长因子建立了直接预估樟子松人工林树冠内一级枝条总数量的回归方程

|
(8) |
式中:Nb为一级枝条总个数,CL为冠长。
2.2 枝条的空间分布 2.2.1 枝条的垂直分布1) 轮枝高度模型 通过简单相关分析,发现轮枝高度与年龄、胸径、树高、轮枝号、枝下高、冠长、立地指数、每公顷株数、林分平均高、林分平均直径均有显著的关系。在此基础上进一步做变量组合和因子筛选分析,建立樟子松人工林轮枝高度模型
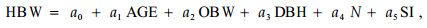
|
(12) |
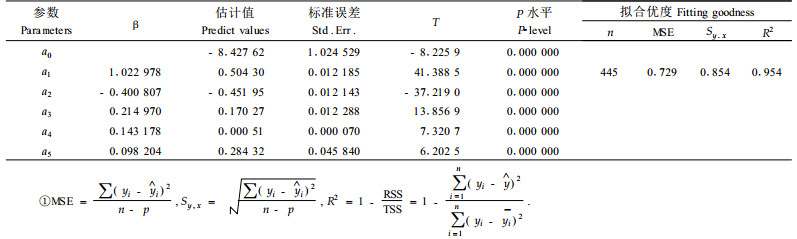
式中:HBW为轮枝高度,AGE为林木年龄,OBW为轮枝序,N为每公顷株数,SI为立地质量,a0、a1, a2, a3, a4, a5均为参数。
参数估计值和拟合统计量见表 2。结果表明:樟子松人工林轮枝高度位置随林分年龄和林木胸径的增大而向上移动,不同林分条件下,立地条件越好、林分密度越大,轮枝高度越高。同一林分中,相同年龄同样大小的林木,同一轮枝高度并没有显著差别。残差图分布没有明显的规律,拟合效果较好(图 1)。
|
|

|
图 1 轮枝高度模型的残差分布图 Fig. 1 Residual for model of estimating whorls height |
2) 枝条垂直分布 在试图应用几种分布函数(Normal、Weibull、Gamma和log Normal)对树冠内的枝条进行拟合后,发现樟子叶松人工林枝条垂直分布并不适合任何一种分布函数。将所有树冠内活枝条根据相对着枝深度绘制折线图(图 2),可以看出,在接近有效冠前(相对着枝深度0.5 ~0.6处),其形状为波浪状,即一“峰"一“谷",说明枝条在垂直方向上的枝条个数呈现一“多"一“少"的趋势,这是因为有效冠内相邻着生的轮枝条其竞争很激烈,无论是对于空间的竞争还是对于光的竞争,致使相邻轮枝枝条个数受到了限制。在有效冠以下,随着着枝深度的增加,枝条个数呈现直线下降趋势。这可能主要是由于树冠上部枝条的摭挡,减弱了下部枝条对光的直接获取能力,自然整枝的结果。

|
图 2 枝条的垂直分布 Fig. 2 Vertical distribution of branches |
3) 每轮枝条的个数 首先应用445轮拟合轮枝样条数目数据与林木因子、林分因子进行简单相关分析,挑选相关系数较大的变量,做逐步回归分析,剔除方程中不显著变量进行简化,重复这一过程,直至能把引起残差自相关的不显著变量进一步剔除为止。建立每轮枝条数目预估模型
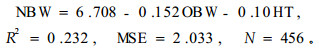
|
(13) |
式中:NBW为每轮枝条数目,HT为林木树高,OBW为轮枝序。
结果表明:每轮枝条的数目与林分立地条件、林分密度等因子无关,与树高和轮枝序呈负相关。但通过方程建立预估每轮枝条的个数效果并不很理想,不论如何变换模型,相关指数均不超过0.25。
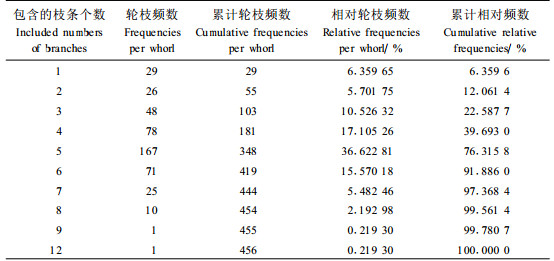
为了进一步描述樟子松人工林树冠轮生枝内枝条数量的分布特性,对样木所有轮枝内的枝条数量进行统计,并计算每轮枝枝条的个数和相对应的频数、相对频数、累计频数以及累计相对频数,结果见表 3。从统计表分析可以看出,所有样木456个轮生枝内的枝条数量从1到12不等(平均值为4.5,标准差为1.6)。在所有轮生枝中,有5个枝条的是最多的,占36.6%。枝条数量在3~6的轮枝数约占80%。各轮生枝个数出现数量“1"基本是在冠顶或冠底,在冠顶是因为极个别梢头出现了分杈或枯死现象,在冠底大部分都出现在最后一轮轮枝。每轮枝条超过9个枝条的情况极少,只占0.44%,本文所调查的456个轮枝中,只有一个轮枝出现9个枝条,另有一个轮枝出现12个枝条这种特殊情况,这2个轮枝均来自z203样地三级木,可能由于这株样木在生长过程中遭受病虫害或其他自然灾害后又重新萌发新的枝条,导致这2轮轮枝枝条数量多于正常轮枝内的数量。
|
|
1) 枝条的方位角分布 本文中枝条方位角数据是以树木生长的北向为0°所测。分别53株解析木数据应用方位角数据进行圆形分布检验,表 4列出了每株树的平均向量长度、平均角及检验统计量。
|
|
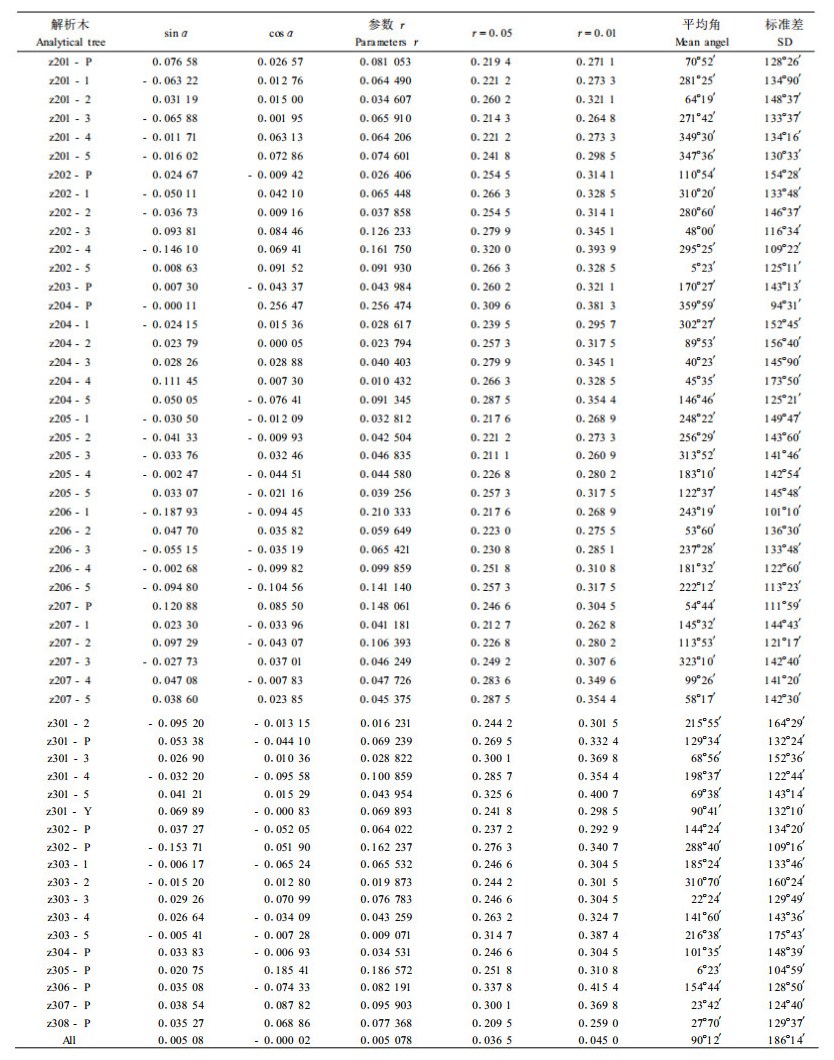
从表 4可以看出,反映集聚程度的r值都很小,接近于0。进一步均匀性检验结果表明全部53株解析木r值均小于表中界值0.05,不存在集中趋势,故方位角的平均角无意义。枝条方位角的分布符合均匀圆形分布。
不同林分条件和不同大小的单株样木枝条方位角的水平分布见图 3a、b,图 3c绘制了全部解析木枝条方位角分布图。

|
图 3 枝条方位角的水平分布图 Fig. 3 Observed branch azimuth distribution a.DBH=21.2 cm,HT=18.5 m,AGE=38 a;b.DBH=15.7 cm,HT=10.75 m,AGE=19 a;c.全部观测数据The entire data set. |
2) 圆-圆相关分析 应用圆-圆相关分析的前提条件是:1)相邻两轮枝具有相同枝条数目;2)每轮枝条的水平分布均符合均匀圆形分布。在53株解析木中共选择每轮包含4个以上枝条数目的相邻轮枝1 42对且每轮枝条水平分布符合均匀圆形分布。每轮中小于4个枝条的成对轮枝没有考虑,因为包含1、2、3个枝条的轮枝由于样本数太少首先无法验证轮枝是否符合均匀圆形分布,另外统计软件DPS7.05也无法计算圆-圆相关分析统计量。因此本文中剔除了包括4个枝条以下的配对轮枝。表 5列出了所有样木匹配的轮枝对数及圆-圆相关分析的统计量。
|
|

从表中可以看出,随着每轮包含的枝条个数增多(从4到7个),圆-圆相关分析相关系数逐渐增大,由于包含8个枝条的轮枝仅有一轮,相关系数统计量有所降低。相邻轮枝圆-圆相关分析结果表明:142对轮枝相关系数均为正相关,其中127对轮枝均通过了H统计量检验,相关显著或极显著(p<0.05或p<0.01)。表 5轮枝对数中,包含4、5个枝条个数的轮枝对数括号内的数字即为没有通过相关系数显著性检验的成对轮枝个数,包含6个枝条以上的轮枝对数中全部通过了H统计量检验。由此进一步说明随着每轮包含枝条个数的增多,相关系数拟合优度越来越好。
进一步分析包含4、5、6个枝条匹配轮枝的平均角度差(由于包含7、8个枝条的样本数较少,没有进行分析)分别为8.9、-3.4和1.5。可以看出随着包含枝条个数的增加,平均角度差的绝对值减少。平均角度差的这种规律可以为建立相邻轮枝位置关系的模型提供基础。一旦确定了具有相同枝条的某一轮枝的位置,通过角度差可以获得它相邻的下一轮枝(或上一轮枝)枝条的方位角。
图 4绘制了单株样木枝条水平分布的俯视图。图中的标记表明枝条水平投影位置,每一圆代表一轮轮枝,最内的圆代表第一轮轮枝,以此类推,最外面的圆代表冠基部最后一轮活轮枝。为了方便比较,绘制的图形仍然以图 3的样木为例。在所有样木中,具有相同枝条轮枝间的正的相关系数都比负的相关系数要大。本文没有考虑相邻轮枝间枝条数目不等的情形,因为无法进行统计量分析。

|
图 4 样木枝条的水平分布 Fig. 4 Observed branch azimuth distribution for selected sample trees a.DBH=21.2 cm, HT=18.5 m, AGE=38 a, n=62; b.DBH=15.7 cm, HT=10.75 m, AGE=19 a, n=52. |
本文应用53株解析木(其中拟合样木48株,检验样木5株)的实测树冠变量因子定量描述了樟子松人工林树冠一级枝条的空间分布。建立的预估模型能较好地预测树冠内总枝条数目。
由于樟子松树木生长的特点,每年只形成一轮轮枝。因此预估一级枝条在树干上的高度位置与预估该轮轮枝在树干上的位置是一致的,本文通过林分和林木变量的筛选组合建立了预估樟子松轮枝的高度预估模型,相关指数R2为0.954,表明应用该模型可以很好地预估一级枝条在树干上的高度位置。但建立预估轮枝内的枝条个数的模型效果似乎并不理想,模型的相关指数很难超过0.25。因此采用了描述性分析樟子松人工林树冠轮生枝内枝条数量的分布状况,所有调查的样木轮生枝内的枝条数量从1到12不等,有5个枝条的轮枝是最多的,占36.6 %。枝条数量为3~6的轮枝数约占80%。各轮生枝个数出现数量“1"基本是在冠顶或冠底,比例较小。
全部53株样木枝条的方位角分布符合均匀圆形分布,因此应用均匀圆形分布可以很好地描述樟子松人工林枝条的水平分布。同时也对具有相同枝条个数的相邻轮枝进行了圆-圆相关分析,结果表明,所有的相邻配对轮枝存在着很强的正相关,并且大部分都通过了显著性检验。
韩兴吉. 1985. 油松树冠枝生长规律的探讨. 北京林学院学报, 7(3): 50-59. |
李凤日. 2004. 长白落叶松人工林树冠形状的模拟. 林业科学, 40(5): 16-24. DOI:10.3321/j.issn:1001-7488.2004.05.003 |
李凤日, 王治富, 王保森. 1996. 落叶松人工林有效冠动态研究(Ⅰ):有效冠的确定. 东北林业大学学报, 24(1): 1-8. |
李俊清, 臧润国, 蒋有绪. 2001. 欧洲水青冈(Fagus sylvatical L.)构筑型与形态多样性研究. 生态学报, 21(1): 151-155. |
刘兆刚, 刘继明, 李凤日, 等. 2005. 樟子松人工林树冠结构的分形分析. 植物研究, 25(4): 465-470. |
马克明, 祖元刚. 2000. 兴安落叶松分枝格局的分形特征. 植物研究, 20(2): 236-240. |
臧润国, 蒋有绪. 1998. 热带树木构筑学研究概述. 林业科学, 34(5): 112-119. DOI:10.3321/j.issn:1001-7488.1998.05.016 |
赵天傍. 1990. 望春玉兰幼龄树体结构规律. 河南林业科技, (2): 16-20. |
祝宁, 陈力. 1995. 植物种群生态学研究现状与进展. 黑龙江科学技术出版社, 69-73.
|
铃木悌司, 大崎惠一, 佐藤创, 等. 1992. コンピュ-タグラフィツクスにょるトドマツ单木の形状表示. 日林志, 74(6): 504-508. |
猪濑光雄. 1982. 单木间の竞合关系にもとづㄑトドマツの成长モデル(第1报)树冠の发达と材积成长量. 林试研报, 318: 103-127. |
Batschelet E. 1981. Circular statistics in biology. London: Academic Press.
|
Carter R E, Miller I M, Klinka K. 1986. Relationships between growth form and stand density in immature Douglas-fir. For Chron, 62: 440-445. DOI:10.5558/tfc62440-5 |
Collin F, Houllier F. 1992. Branchiness of Norway spruce in northeastern France: predicting the main crown characteristics from usual tree measurements. Ann Sci For, 48: 511-538. |
Doruska P F, Burkhart H E. 1994. Modeling the diameter and locational distribution of branches within the crown of loblolly pine trees in unthinned plantation. Can J For Res, 24: 2362-2376. DOI:10.1139/x94-305 |
Hashimoto R. 1990. Analysis of the morphology and structure of crowns in a young sugi stand. Tree Physiol, 6: 119-134. DOI:10.1093/treephys/6.2.119 |
Hashimoto R. 1991. Canopy development in young sugi stands in relation to changes with age in crown morphology and structure. Tree Physiol, 8: 129-143. DOI:10.1093/treephys/8.2.129 |
Li Fengri, Jia Weiwei. 2004. Modeling locational distribution of branches within the crowns of Larix olgensis trees in China. Jour Korean For Soc, 93(6): 335-343. |
Maguire D A, Kershaw J A, Hann D W. 1991. Predicting the effects of silvicultural regime on branch size and crown wood core in Douglas-fir. For Sci, 37: 1409-1428. |
Maguire D A, Moeur M, Bennett W S. 1990. Simulating branch diameter and branch distribution//Burkhart H E. Research in forest mensuration growth and yield. Virgina Polytechnic Institute and State University School of Forestry and Wildlife Resources Publ FWS-2-90, 85-94
|
Mardia K V. 1972. Statistics of direction data. London: Academic Press.
|
Tombleson J D, Grace J C, lnglis C S. 1990. Response of radiata pine branch characteristics to site and stocking//James R N, Tarlton G L. New approaches to spacing and thinning in plantation forestry. N Z Fr Serv FRI Bull, 151: 229-231. |