 2007, Vol. 43
2007, Vol. 43文章信息
- 乌云塔娜, 谭晓风, 李秀根, 张林, 邓建军.
- Wuyun Tana, Tan Xiaofeng, Li Xiugen, Zhang Lin, Deng Jianjun.
- 13个‘新世纪’梨后代品种S基因型的鉴定
- Identification of S-Genotypes of 13 Progenies of 'Shinseiki' Pear
- 林业科学, 2007, 43(9): 116-122.
- Scientia Silvae Sinicae, 2007, 43(9): 116-122.
-
文章历史
- 收稿日期:2006-07-18
-
作者相关文章



2. 中国农业科学院郑州果树研究所 郑州 450009
2. Zhengzhou Fruit Institute, Chinese Academy of Agriculture Sciences Zhengzhou 450009
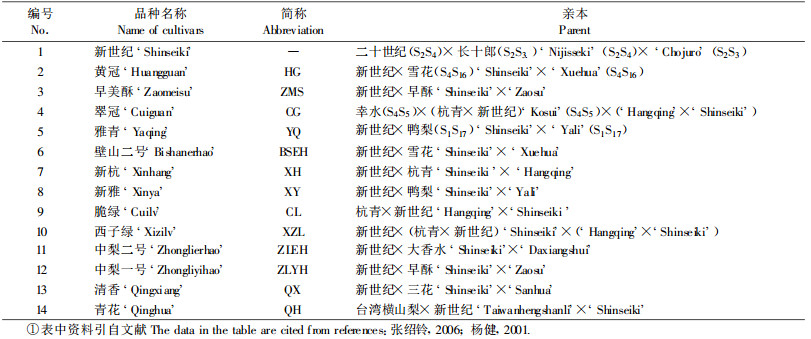
‘新世纪’(Shinseiki)梨是日本培育的沙梨(Pyrus pyrifolia)的优良品种(梁林平等, 2001; 杨健等, 2001)。近年来中国、日本、韩国利用‘新世纪’梨进行杂交选育了许多优良品种,并广泛地栽培利用,这些品种都属于沙梨系统。沙梨是配子体自交不亲和品种,生产上常采用配置授粉品种或人工授粉等方法来克服自交不亲和性(佐藤昭宏, 2001)。自交不亲和梨品种无论配置授粉品种或人工授粉,均要考虑品种间的亲和性。而品种间的亲和性和S基因型间存在密切联系,即S基因型相同,则为自交不亲和,否则为自交亲和(Saito et al., 1999; Sassa et al., 1993; Ishimizu et al., 1996)。因此,梨品种S基因型的鉴定,对授粉品种配置、人工授粉以及梨的遗传改良及栽培利用具有一定的指导意义。国内外对梨自交不亲和性方面进行了大量的研究,目前已分离鉴定了35个S等位基因,确定了80多个品种的S基因型(谭晓风等, 2005a; 2005b;乌云塔娜等, 2005; 2006a; 2006b; Saito et al., 1999; Sassa et al., 1993; 1997;Ishimizu et al., 1996; 1999a; 1999b; Norioka et al., 1995; 1996; Shin et al., 1995; Kim et al., 2002; Ushijima et al., 1998)。日本通过分子生物学方法已经确定‘新世纪’梨的S基因型,为S3S4(佐藤昭宏, 2001)。梨的自交不亲和性由单基因位点控制,遵循孟德尔分离规律,故‘新世纪’梨的后代品种应该含有S3基因和S4基因中的一个。因此,本文选用13个‘新世纪’梨后代品种,分别用S3和S4基因特异内切酶切割,确定各品种的一个S基因,而另一个S基因通过PCR片段的回收测序来确定。这些品种S基因型的确定为沙梨授粉品种配置及人工授粉提供重要的理论依据。
1 材料与方法 1.1 试验材料1) 植物材料 ‘新世纪’、‘黄冠’等14个梨品种叶片采集于郑州果树研究所,样品及亲本见表 1。采集叶片2 g左右,保存于-80 ℃备用。
|
|
2) PCR引物 梨S基因由5个保守区、1个高变区及1段内含子组成。梨S等位基因的多态性主要来自于高变区(HV)的变异,通常根据高变区的不同来确认不同的S等位基因。引物设计在S基因高变区周边(图 1)。引物由上海基康有限公司合成。引物序列分别为:P1(FTQQYQ): 5′-TTTACGCAGCAATATCAG-3′;P2(IIWPNVV): 5′-AC(A/G)TTCGGCCAAATAATT-3′。

|
图 1 梨S基因结构及引物位置 Fig. 1 The structure and primer position of pear S gene SP:信号肽Signal peptide; HV:高变区Hyper variable region; C1-5:保守区Conserved; 空格:外显子Exson. |
3) 药品试剂 PCR相关试剂购自大连宝生物技术有限公司,限制酶购自上海生物技术有限公司。
1.2 试验方法1) 梨叶DNA的提取、纯化,纯度和浓度测定 梨叶DNA的提取采用CTAB法(乌云塔娜等, 2003),DNA纯化采用饱和酚纯化法(乌云塔娜等,2003),纯度和浓度测定采用DU-640核酸蛋白分析仪。
2) PCR扩增反应体系和循环条件 PCR扩增反应体系和循环条件与谭晓风等(2005b)基本相同。
3) 酶切检测 选用‘新世纪’梨的杂交优良后代品种。‘新世纪’梨的S基因型为S3S4(佐藤昭宏, 2001),按照孟德尔分离规律,‘新世纪’梨的后代一定含有S3或S4等位基因。首先利用S4等位基因特异性限制酶来切割所有品种PCR产物,根据是否被切割和酶切片断大小来确定哪些品种含有S 4等位基因。对不含S4等位基因的梨品种PCR产物,再用S3等位基因的特异性限制酶来切割,并根据是否被切割及切割片段大小来确定是否含有S3等位基因。S3和S4等位基因的特异性限制酶切割片段大小见表 2。限制酶反应体系按说明书进行。产物检测用2.0%的琼脂糖凝胶电泳进行检测,以100 bp的DNA ladder作分子质量参照物。
|
|

4) DNA序列测定 根据酶切鉴定13个梨品种的1个S等位基因后,另一个S等位基因通过DNA序列测定及生物信息学分析来确定。DNA序列测定时,首先用高保真酶PCR扩增供试品种的基因组DNA,并用S3等位基因或S4等位基因特异性限制酶来切割PCR扩增产物,回收未被酶切的370 bp左右(另一个S等位基因)的DNA片段进行纯化、DNA测序,并利用生物信息学软件进行相似性比较。DNA测序委托上海基康生物技术公司完成。
2 结果与分析 2.1 S基因特异引物PCR检测及酶切检测供试的14个梨品种基因组DNA进行S基因特异性引物PCR扩增结果表明,14个梨品种均扩增出1条370 bp左右的DNA扩增片段,难以分离2个S等位基因(图 2)。

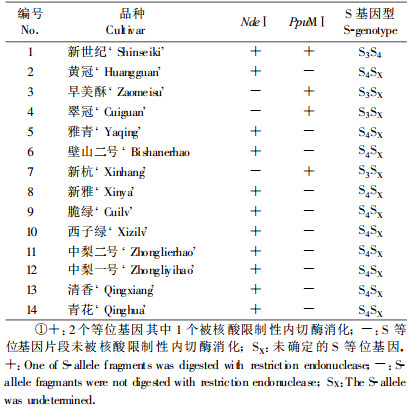
由于‘新世纪’梨的S基因型为S3S4,供试品种均为‘新世纪’梨的后代,因此这些‘新世纪’梨的后代品种中含有S3等位基因或S4等位基因,因此首先利用S4等位基因特异酶NdeⅠ切割供试的14个梨品种S基因特异PCR产物。结果表明:黄冠、雅青、壁山二号、新雅、脆绿、西子绿、中梨二号、中梨一号、清香、青花10个品种PCR产物被NdeⅠ切割,且酶切产物大小与‘新世纪’梨S基因产物被NdeⅠ切割情况一致,大小在140 bp左右和228 bp左右(图 3),因此可确定以上10个品种含有S4等位基因。

|
图 3 NdeⅠ酶切图 Fig. 3 Restriction pattern of NdeⅠ |
另外3个品种新杭、翠冠、早美酥未被NdeⅠ切割,可能含有S3等位基因,因此将这3个品种的PCR产物,用S3等位基因的特异性限制酶PpuMⅠ进行切割。结果表明新杭、翠冠、早美酥均被 PpuMⅠ切割,且酶切产物大小与‘新世纪’梨S基因PCR产物被Ppu MⅠ切割情况一致(图 4),大小在107 bp和270 bp左右,因此可以确定新杭、翠冠、早美酥含有一个S3等位基因。

通过NdeⅠ和PpuMⅠ切割,可以确定黄冠等13个品种的1个S等位基因(表 3)。
|
|
DNA序列分析结果表明:用P1和P2可扩增出S基因保守区1和2、HV区、内含子、2条引物序列及外显子序列(图 5、图 6)。序列相似性比较表明:黄冠、翠冠、早美酥、雅青、壁山二号、新杭、脆绿、西子绿、中梨二号、中梨一号、清香、新雅、青花的另1条S等位基因产物分别与S16、S5、S35、S17、S16、S1、S3、S1、S31、S35、S7、S17、S1等位基因相似性达100%(图 5、图 6)。因此,它们分别含有S16、S5、S35、S17、S16、S1、S 3、S1、S31、S35、S7、S17、S1等位基因。

|
图 5 S基因的DNA序列比较 Fig. 5 DNA sequence alignment of S-genes HG、BSEH为品种简称,见表 1。下划线:P1,也是保守区1;双下划线:半胱氨酸残基;粗线:保守区2;虚下划线:HV区。HG and BSEH are No. of cultivars, see Tab. 1. Underline:P1,conserved region 1;Crewel:Cys;Heavy line:Conserved region 2;Dashed underline: HV region. S16:A Y249431;S5:AB045711;S35:DQ224344;S17:AY249432;S1:DQ515793;S3:AB025421;S7:AB002143;S31:DQ072113. |

|
图 6 S基因的DNA序列比较(续) Fig. 6 DNA sequence alignment of S-genes(continued) HG、BSEH为品种简称,见表 1。虚下划线:HV区;点式下划线:内含子;波浪线:P2。HG and BSEH are No. of cultivars, see Tab. 1. Dashed underline:HV region; Point underlined: Intron; Cutting plane line:P2. S16:A Y249431;S5:AB045711;S35:DQ224344;S17:AY249432;S1:DQ515793;S3:AB025421; S7:AB002143;S31:DQ072113. |
通过分子生物学和生物信息学方法鉴定了‘新世纪’梨后代13个优良品种的S基因型,黄冠、翠冠、早美酥、雅青、壁山二号、新杭、脆绿、西子绿、中梨二号、中梨一号、清香、新雅、青花等品种的S基因型分别为S4S16、S3S 5、S3S35、S4S17、S4S16、S1S3、S3S4、S1S4、S4S31、S4S35、S 4S7、S4S17、S1S4。
本文选用的13个梨品种均为‘新世纪’梨的杂交优良后代。梨自交不亲和性是属于配子体自交不亲和性,其特点为被单个S位点的复等位基因所控制,遗传模式上遵循孟德尔分离规律。‘新世纪’梨的S基因型为S3S4,因此以‘新世纪’梨为亲本进行杂交后获得的后代一定含有S3或S4等位基因。故本文利用‘新世纪’梨的S基因型,与它具有亲缘关系的梨品种为试验材料,首先确定是否含有S4等位基因(NdeⅠ比PpuMⅠ便宜),结果表明黄冠、雅青、壁山二号、新雅、脆绿、西子绿、中梨二号、中梨一号、清香、青花10个品种含有S4等位基因。然后对不含S4等位基因的梨品种新杭、翠冠、早美酥,用S3等位基因的特异性内切酶来切割,结果表明这3个品种含有S3等位基因。13个梨品种的另外1条等位基因回收测序后在GenBank中进行Blast,结果表明,测序的序列均可以在GenBank中找到,这样可以成功地鉴定13个梨品种的S基因型。
13个梨品种的亲本基因型有的已被确定,如雪花(S4S1 6)、幸水(S4S5)、鸭梨(S1S17),而其他亲本品种的S基因型的研究鲜见报道,故难以通过亲缘关系来进一步验证这13个梨品种的S基因型。但可以利用本研究结果推测出亲本的S等位基因,如早美酥S基因型为S3S35,亲本为‘新世纪’(S3S4)×早酥,按照孟德尔遗传规律早美酥的S3等位基因来自于‘新世纪’梨,而另一个等位基因S35来自于早酥,故早酥一定含有S35等位基因;中梨一号(S4S35)的S基因型也证明了这一点。中梨二号S基因型为S4S31,其中S4等位基因来自于‘新世纪’梨,S31等位基因来自于大香水,故大香水应该含有1个S31等位基因;同理可以推测三花含有1个S7等位基因,台湾横山梨含有1个S1等位基因。从新杭(S1S3)可以推测出杭青含S1等位基因,从脆绿(S3S4)推测出杭青的另一个S等位基因可能为S3或S4。
梁林平, 王永恒, 仲卫, 等. 2001. 中早熟日本梨新品种——新世纪. 西北园艺, (4): 43. |
谭晓风, 乌云塔娜, 中西 テッ, 等. 2005a. 中国梨品种自交不亲和基因的分离鉴定. 中南林学院学报, 25(1): 1-3. |
谭晓风, 乌云塔娜, 中西 テッ, 等. 2005b. 中国沙梨7个自交不亲和新基因的分离与测序. 中南林学院学报, 25(4): 1-6. |
乌云塔娜, 谭晓风, 曹玉芬, 等. 2006a. 白梨新S基因的克隆. 园艺学报, 33(2): 385-388. |
乌云塔娜, 谭晓风, 李秀根, 等. 2006b. 梨自交不亲和新基因S12-RNase的分离鉴定及序列分析. 林业科学, 42(2): 117-121. |
乌云塔娜, 谭晓风, 毕方铖, 等. 2005. 7个白梨品种S基因型确定和2个新S基因的鉴定. 中南林学院学报, 25(4): 7-12. DOI:10.3969/j.issn.1673-923X.2005.04.002 |
乌云塔娜, 张党权, 谭晓风. 2003. 梨不同DNA提取方法的效果研究. 中国生物工程杂志, 23(7): 98-101. |
杨健, 李秀根. 2001. 梨优良种质资源及其在杂交育种上的利用. 中国种业, (5): 29-30. DOI:10.3969/j.issn.1671-895X.2001.05.022 |
张绍铃, 黄绍西, 吴俊. 2006. 日本梨优良品种资源在梨育种上的应用. 中国南方果树, 35(5): 48-49. DOI:10.3969/j.issn.1007-1431.2006.05.024 |
佐藤昭宏. 2001.RT-PCRによるセィョナシS遗传子型の推定.修士学位论文(日本)
|
Kim H T, Hirata Y, Nou I S, et al. 2002. Determination of S-genotypes of pear(Pyrus pyrifolia)cultivars by S-RNase sequencing and PCR-RFLP analyses. Mol Cell, 13(3): 444-451. |
Ishimizu T, Sato Y, Saito T, et al. 1996. Identification and partial amino acid sequenes of seven S-RNase associated with self-incompatibility of Japanese pear (Pyrus pyrifolia). J Biocbem, 120: 326-334. |
Ishimizu T, Inoue K, Shimonaka M, et al. 1999. PCR-based method for identifying the S-genotypes of Japanese pear cultivars. Theor Appl Genel, 98: 961-967. DOI:10.1007/s001220051156 |
Ishimizu T, Mitsukami Y, Shinkawa T, et al. 1999. Presence of asparagines-linked N-acetylglucosamine and chitobiose in Pyrus serotina S-RNase associated with gamaetophytic self-incompatibility. Eur Biochem, 263: 624-634. DOI:10.1046/j.1432-1327.1999.00499.x |
Norioka N, Norilka S, Ohnishi Y, et al. 1996. Molecular cloning and nucleotide sequenece of cDNA encoding S-allele specific stylar RNases in a self-incompatible cultivar and its self-compatible mutant of Japanese pear. J Biochem, 120: 335-345. DOI:10.1093/oxfordjournals.jbchem.a021418 |
Norioka N, Ohnishi Y, Norioka S, et al. 1995. Nucleotide sequences of cDNAs encoding S2- and S4-RNases (D49527 and D49528 for EMBL) from Japanese pear(Pyrus pyrifolia Nakai). Plant Physiol, 108: 1343-1347. |
Saito T, Terai O, Norilka S. 1999. PCR-based method for identifying the S-genotypes of Japanese pear cultivars. Theor Appl Genel, 98: 961-967. DOI:10.1007/s001220051156 |
Sassa H, Hirano H, Ikehashi H, et al. 1993. Identification and characterization of stylar glycoproteins associated with self-incompatibility genes of Japanese pear. Mol Gen Gent, 241: 17-25. |
Sassa H, Hirano H, Nishio T, et al. 1997. Style-specific self-compatible mutation caused by deletion of the S-RNase gene in Japanese pear(Pyrus serotina). Plant J, 12: 223-227. DOI:10.1046/j.1365-313X.1997.12010223.x |
Shin H, Yoshiji O. 1995. Stylar basic proteins corresponding to 5 self-incompatibility alleles of Janpanese pears. J Japan Soc Hort Sci, 64(3): 471-478. DOI:10.2503/jjshs.64.471 |
Ushijima K, Sassa H, Hirano H, et al. 1998. Characterization of the flanking regions of the S-RNase genes of Japanese pear (Pyrus serotina) and apple (Malus× domestica). Gene, 211(1): 159-167. DOI:10.1016/S0378-1119(98)00105-X |