 2007, Vol. 43
2007, Vol. 43文章信息
- 王旭丽, 臧睿, 王磊, 康振生, 黄丽丽.
- Wang Xuli, Zang Rui, Wang Lei, Kang Zhensheng, Huang Lili.
- 苹果树Valsa malicola的发现及其致病性研究
- The Occurrence of Valsa malicola on Apple Trees and Its Pathogenicity
- 林业科学, 2007, 43(9): 23-26.
- Scientia Silvae Sinicae, 2007, 43(9): 23-26.
-
文章历史
- 收稿日期:2007-01-09
-
作者相关文章



由Valsa spp.引起的苹果树(Malus pumila)腐烂病在我国发生普遍、危害严重,严重制约了我国苹果产业的发展和农民增收,也严重影响了苹果的对外出口。据陕西省等苹果主产区的统计,苹果树腐烂病病株率目前已经高达70%,病园率达100%,成为影响苹果生产的主要病害。臧睿等(2007)对苹果树腐烂病菌的种类鉴定、防治理论方面进行了系统研究,本文则报道了分离自陕西苹果树腐烂病病斑上的几株分离物的形态学和rDNA-ITS核苷酸序列比对鉴定结果,并研究了其对离体枝条的致病性特征。
1 材料与方法 1.1 供试菌种的采集、分离在陕西省岐山县周公庙乡念郭村采集典型症状的苹果树腐烂病病皮,按常规组织块分离法从病斑的病健交界处分离,所得2株单孢分离株(标号03-1和03-7)分别保存在PDA斜面上(4 ℃)备用。采自陕西省富县交道乡苹果枯枝上子实体样本,经75%乙醇表面消毒后直接挑取子实体进行分离,共获得了3个分离株(标号SXFX-V2、SXFX-V4和SXFX-V6),分别保存在PDA斜面上(4 ℃)备用。
1.2 形态学 1.2.1 有性型的鉴定用双面刀片直接切取田间苹果树皮上所产生的子囊座,分别在显微镜(LEICA DM LB2)和实体解剖镜(LEICA S6D)下观察其外部形态及纵横切面的形态特征。
1.2.2 无性型的鉴定供试菌种接种离体苹果枝条,待其形成产孢体后(臧睿,2006),在显微镜(LEICA DM LB2)和实体解剖镜(LEICA S6D)下观察其外部形态及纵横切面的形态特征。
1.3 培养特性 1.3.1 菌落颜色将供试菌种接于PDA平板,25 ℃黑暗条件下培养,从第2天开始每天观察记录菌落的颜色、形状,气生菌丝的疏密程度。
1.3.2 产孢体特征将供试菌种接于PDA平板,25 ℃黑暗条件下培养3 d后用打孔器(Φ=5 mm)在菌落边缘取菌饼分别接种于PDA平板和20%ABA(臧睿,2006)平板中央(15 mL培养基)。每分离株设3重复皿,置于25 ℃黑暗条件下培养,每天观察并记录产孢体形成的初始时间、个数、大小范围及着生方式(单生或聚生)。
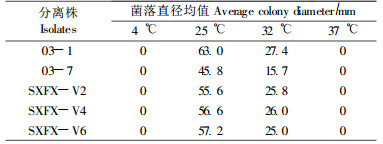
1.4 对温度的适应性将供试菌种接于PDA平板,置不同温度条件下(4、25、32、37 ℃)暗培养3 d后测量菌落直径(Adams et al., 2005),每处理3个重复皿。
1.5 分子鉴定 1.5.1 DNA提取供试菌株经PD(马铃薯200 g,葡萄糖20 g,水1 000 mL)液体培养5~6 d,收集菌丝体进行冷冻干燥,后保存于-70 ℃备用。采用Lee等(1990)的方法提取菌株的基因组总DNA。
1.5.2 目标DNA的PCR扩增和核苷酸序列的测定采用引物ITS1和ITS4(White et al., 1990)扩增rDNA的ITS1+5.8+ ITS2片段。引物序列为ITS1:5'TCCGTAGGTGAACCTGCG 3'、ITS4:5'TCCTCCGCTTATTGATAT 3'。PCR反应体系为25 μL,反应条件为:94 ℃预变性2 min;94 ℃变性30 s,51 ℃退火30 s,72 ℃延伸40 s, 共38个循环,最后72 ℃延伸5 min。扩增产物(3 μL)经1.5%的琼脂糖电泳检测后,其余用QIAquick PCR产物纯化试剂盒(Qiagen公司)进行回收。
纯化PCR产物适当稀释后用BigDye3.1测序试剂盒进行测序,测序引物用ITS1,反应条件为:95 ℃预变性2 min;95 ℃变性10 s,60 ℃延伸4 min,共进行35个循环,反应体积为10 μL。测序产物纯化后在ABI 3130XL序列测定仪器进行序列测定。
1.5.3 序列比对和分析采用ClustalX1.8 (Thompson et al., 1997)对序列片段进行比对,运用MEGA 3.1 (Kumar et al., 2004)进行序列分析,分别采用邻接法和最大简约法构建系统发育树。对所构建的系统发育树进行自举检验(重复1 000次),以获得分支的支持率。最近的研究表明杨树腐烂病菌V. sordida与V. malicola的系统发育关系极近(Admas et al., 2006),因此本文对采自陕西杨凌的杨树腐烂病菌进行序列测定并一起进行系统发育分析。在分析过程中选择Diaporthe vaccinii为外群。
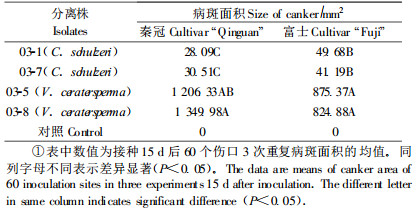
1.6 致病性研究分别用供试分离株和V. ceratosperma的2个分离株03-5和03-8接种2年生健康的“秦冠”和“红富士”苹果枝条,以菌饼(Φ 7 mm)作为接种体进行接种(臧睿,2006)。每个枝条上5个接种点,每个分离株接5个枝条,每个枝条上随机选取一个伤口,以无菌PDA圆饼(Φ 7 mm)作为接种对照。试验重复3次。接种后的枝条插入湿沙盘置25 ℃培养,保湿48 h,每天测量病斑的长度和宽度,并按椭圆面积计算病斑面积,并在接种后15 d对处理间的差异进行Duncan多重范围检验。
2 结果与分析 2.1 形态学研究 2.1.1 有性型的形态特征富县苹果枯枝上的子囊座形态特征为:子囊座像截短的圆锥形或圆顶状、扁平,至少由11个子囊壳组成且排成一圈或在邻近于木头的子座底部不规则的排列(图版Ⅰ-1)。子囊座盘(图版Ⅰ-2)圆形或椭圆形,孔口沿着边缘排列成环形,而中央区域仍为寄主组织(图版Ⅰ-2)。有时孔口和子囊座盘一样高,有时颈伸出子囊座盘外,且向外弯曲(图版Ⅰ-3);子囊壳球形,具长颈(图版Ⅰ-4)。子囊棍棒状,大多数含8个孢子,极少数含4~6个孢子,大小为(37~50)μm×(7.5~12.5) μm(图版Ⅰ-5);子囊孢子无色,稍弯曲,有时不匀称,大小为(11~14) μm×(2.2~2.5) μm(图版Ⅰ-6)。根据以上形态特征将其鉴定为V. malicola Z.Urb。

|
图版Ⅰ 苹果树Valsa malicola的发现及其致病性研究 Plate Ⅰ The occurrence of Valsa malicola on apple trees and its pathogenicity 1~6:分离株SXFX-V2的形态学特征 1.子囊座;2.子囊座盘和孔口;3.子囊座的纵切面;4.子囊座的横切面;5.子囊;6.子囊孢子。7~14:分离株03-7的形态学特征 7.产孢体座;8,9.产孢体的纵横切面:有一个腔(包含多个室);10.分生孢子;11,12. PDA上分离株03-1的菌落形态正面和反面观;13.分离株03-1在20%ABA上形成的产孢体正面观(右下角为产孢体溢出黄褐色的胶质体);14.在20%ABA上形成的产孢体反面观。 1~6:Morphologic characteristics of the isolates of SXFX-V2 1. Ascostromata. 2. Disc and and ostioles; 3. Longitudinal section of ascostroma; 4. Traverse section of ascostroma; 5. Asci; 6. Ascospores. 7~14:Morphologic characteristics of the isolates of 03-7 7. Conidiomata stromata; 8, 9. Longitudinal and traverse section of conidioma showing a single locule composed of numerous chambers; 10. Allantiod conidia; 11, 12. The surface and reverse of the colony morphology on PDA; 13. The surface of conidiomata formed on 20%ABA(Insert shows brown cirrhi exuded by conidioma); 14. The reverse of conidiomata formed on 20%ABA. |
将03-1和03-7分离株接种离体苹果枝条,8~9 d就可形成产孢体。产孢体特征为:子座圆形或圆锥形;产孢体盘为单孔口(图版Ⅰ-7);产孢体为单腔室,但包含无数不规则但同中心的室(图版Ⅰ-8,9);分生孢子香蕉形,大小为(6.25~7.5)μm×(1.25~1.4)μm(图版Ⅰ-10)。依据形态特征将其归为Cytospora schulzeri Sacc. & P.Syd。
2.2 培养特性5个分离株在PDA上的菌落初为白色,后期中央乳白色,外围白色,呈放射状,气生菌丝稀少,背面为乳黄色。在观测期内(30 d)未见产孢体的形成,而在20%ABA培养基上第6天时就可产生大量(140~250个/皿)微小(0.1~0.5 mm)的产孢体,且有黄色的胶质体流出(图版Ⅰ-11~14)。
2.3 对温度的适应性表 1说明:各分离株在37 ℃高温下不能生长的特性可区别于Valsa属中的高温生长种类。尽管各分离株在25 ℃和32 ℃培养条件下生长良好,但菌落直径显示其生长速率有明显差异,4 ℃下也能够缓慢生长,10 d时菌落直径可达5 mm。
|
|
本试验所测各分离株最后序列长度为481 bp,5条序列中4条完全相同,另外一条(SXFX-V4)仅在一个位点发生了G/C的颠换。其rDNA的ITS(含5.8S基因)区段与美国和南非的V. malicola序列差异为0.98%(K2P距离)(Kimura,1980)。V. malicola与V. sordida间的平均遗传距离为3.39%。ITS序列聚类结果表明,陕西各分离株与美国和南非V. malicola被聚为一类(图 1),用2种方法(NJ/MP)构建的系统发育树的拓扑结构完全一致,自举支持率很高(>90%)。

|
图 1 用ITS序列构建的系统发育树 Fig. 1 Phylogram using ITS sequences data 分支上值为NJ/MP自举支持率,1 000次重复;SXYLyf为采自陕西杨凌杨树腐烂病的分离株编号;DQ243802和DQ243792分别为美国和南非苹果树上的V. malicola的Genbank号。Numbers at the nodes refer to bootstrap support given as percentage from pseudo 1 000 replicates by NJ/MP;SXYLyf was the isolate from Populus Valsa canker in Yangling,Shaanxi;DQ243802 and DQ243792 were V. malicola of apple tree in USA and South Africa. |
V. malicola的5个分离株接种“秦冠”和“富士”2个品种的离体枝条,可形成黄褐色水浸状的凹陷,但扩展缓慢,面积很小,并在8~9 d后产生大量产孢体(表 2)。而苹果树腐烂病菌的常见种V. ceratersperma的2个分离株03-5和03-8形成的病斑面积很大,在接种后20 d才能在接种枝条上形成产孢体(表 2)。
|
|
V. malicola Z. Urb.和C. schulzeri Sacc. & P. Syd.最早分别发现于1956年和1899年,主要寄生于苹果属的植物。Hayova等(1998)曾提及该种在中国可能有分布,但并未报道样本来源等详细信息。本研究证实了V. malicola在陕西的分布:从全国150个分离株中发现了5个来源于陕西富县(有性型)和岐山县(无性型)的分离株为V. malicola和C. schulzeri,但在其他地区的分布情况还需进一步研究。
Adams等(2005)在鉴定桉树(Eucaluptus sp.)腐烂病菌时发现,Valsa属20个种中有4个种(V. brevispor, V. eugeniae, V. fabian, C. eucalyptina)可以在37 ℃高温下生长而其他各种则不能。因此,他们提出在鉴定Valsa属下各种时应将其能否在37 ℃下生长作为鉴定的辅助依据。本试验研究了陕西V. malicola的各分离株在37 ℃下的生长情况,发现其不能生长,因此,在以后鉴定该种时要验证此性状。
本研究发现V. malicola在20% ABA培养基上形成的产孢体较为独特,与苹果树上其他种的产孢体区别较大,可将在20% ABA产孢特点作为鉴定V. malicola的辅助依据。另外,由于Cytospora属下各个种在自然条件下与在人工培养条件下的形态特征有相当大地变化,同时在自然界有性型非常稀少且在培养条件下不易产生(Adams et al., 2005)。因此,依据形态特征作出准确鉴定是非常困难的,而DNA序列分析与形态学鉴定方法相结合可以大大提高鉴定的准确性。
由于ITS区进化速度相对较快,可用于物种的区别鉴定,也用于种群遗传分化研究(White et al., 1990)。Adams等(2002;2005;2006)利用该区段序列分别鉴定了由Valsa属引起的核果类果树溃疡病菌;而且Evertt等(2006)还应用该技术对柑橘(Citrus reticulata)黑斑病菌进行了重新分类和鉴定,将Guignardia citricarpa更正为G. mangiferae。另外,也可通过18S、5.8S和ITS序列区分禾顶囊壳的3个变种,还可以进一步区分禾顶囊壳小麦(Triticum aestivum)变种能否侵染黑麦(Secale cereale)的分离物,揭示顶囊壳属-瓶梗霉属复合群的关系(Bryan et al., 1995)。基于序列分析的分子鉴定手段将大大提高植物病原真菌鉴定的准确性和可靠性。
关于V. malicola对苹果的致病性,Proffer等(1989)曾将其接种于经冰冻处理的苹果幼苗和成株,发现不能侵入成株的冻伤部位及健康组织,而只定植于幼苗受冻伤的部位,因此,认为该种类不是致病菌。但Adams等(2006)在鉴定南非树木上腐烂病的病原菌种类时却将V. malicola定为致病菌。本研究表明V. malicola有一定的弱致病性。关于其在田间的致病性有待进一步研究。
王磊, 臧睿, 黄丽丽, 等. 2005. 陕西省关中地区苹果树腐烂病调查初报. 西北农林科技大学学报:自然科学版, 33(增): 98-100. |
臧睿. 2006.陕西苹果树腐烂病菌不同分离株生物学特性及致病性研究.西北农林科技大学学位论文 http://cdmd.cnki.com.cn/Article/CDMD-10712-2006179875.htm
|
臧睿, 黄丽丽. 2007. 苹果树腐烂病菌分生孢子萌发及其影响条件研究. 西北农业学报, 16(1): 64-67. DOI:10.3969/j.issn.1004-1389.2007.01.014 |
Adams G C, Surve-Iyer R S, Iezzoni A F. 2002. Ribsomal DNA sequence divergence and group I introns within the Leucostoma species L. cinctum L. persoonii and L. parapersoonii sp. Nov., Ascomycetes that cause Cytospora canker of fruit trees. Mycologia, 94(6): 947-967. DOI:10.1080/15572536.2003.11833153 |
Adams G C, Wingfield M J, Common R, et al. 2005. Phylogenetic relationships and morphology of Cytospora species and related teleomorphs (Ascomycota, Diaporthales, Valsaceae) from Eucalyptus. Studies in Mycology, 52: 1-142. |
Adams G C, Roux J, Wingfield M J. 2006. Cytospora species (Ascomycot, Diaporthales, Valsacea): introduced and native pathogens of trees in South Africa. Australasian Plant Pathology, 35: 521-548. DOI:10.1071/AP06058 |
Bryan G T, Daniels M J, Osbourn A E. 1995. Comparison of fungi within the Gaeumannomyces-Phialophora complex by analysis of ribosomal DNA sequences. Applied and Environmental Microbiology, 61: 681-689. |
Evertt K R, Rees-George J. 2006. Reclassification of an isolate of Guignardia citricarpa from New Zealand as Guignardia mangiferae by sequence analysis. Plant Pathology, 55: 194-199. DOI:10.1111/j.1365-3059.2006.01334.x |
Hayova V P, Minter D W. 1998. Valsa malicola. IMI Descriptions of fungi and bacteria, No.1366. CAB International, U.K.
|
Kimura M. 1980. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution, 15: 111-120. |
Kumar S, Tamura K, Nei M. 2004. MEGA3:Integrated software for molecular evolutionary genetics analysis and sequence alignment. Briefings in Bioinformatics, 5: 150-163. DOI:10.1093/bib/5.2.150 |
Lee S B, Taylor J W. 1990. Isolation of DNA from fungal mycelia and single spores//Innis M A, Gelfand D H, Sninsky J J, et al. PCR protocols, a guide to methods and applications. San Diego, California: Academid Press, 282-287
|
Proffer T J, Jones A L. 1989. A new canker disease of apple caused by Leucostoma cincta and other fungi associated with cankers on apple in Michigan. Plant Disease, 73: 508-514. DOI:10.1094/PD-73-0508 |
Thompson J D, Gibson T J, Plewniak F, et al. 1997. Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research, 24: 4876-4882. |
White T J, Bruns T D, Lee S B, et al. 1990. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics//Innis M A, Gelfand D H, Sninsky J J, et al. PCR protocols, a guide to methods and applications. San Diego, California: Academic Press, 315-322
|