 2007, Vol. 43
2007, Vol. 43文章信息
- 龙翠玲, 余世孝.
- Long Cuiling, Yu Shixiao.
- 茂兰喀斯特森林的林隙物种组成动态及更新模式
- Dynamics of Species Composition and Generation Pattern in the Gaps of Karst Forest in Maolan, Guizhou Province
- 林业科学, 2007, 43(9): 7-12.
- Scientia Silvae Sinicae, 2007, 43(9): 7-12.
-
文章历史
- 收稿日期:2006-06-19
-
作者相关文章



2. 贵州师范大学地理与生物科学学院 贵阳 550001
2. School of Geography and Life Science, Guizhou Normal University Guiyang 550001
喀斯特森林生态环境特殊,蕴藏着丰富的生物多样性,群落结构动态及维持机制的研究是喀斯特森林生态研究的重要内容之一。从林隙更新的角度探讨群落结构动态及维持机制日益成为国内外生态学研究的热点(臧润国等,1999; 王周平等, 2001; 熊小刚等, 2002; 何永涛等, 2003;Arnaga, 1988; Whitemore, 1989; Lertzman, 1992)。有关喀斯特森林的植被类型、群落学特点及更新等方面已有一些研究(朱守谦,1993;刘济明,2000),但喀斯特森林林隙物种组成动态及更新模式的研究未见报道,对其群落结构动态及维持机制知之甚少。迄今为止,关于林隙更新的研究已较为充分,但林隙物种组成结构及种间关系随林隙发育的动态变化及树种更新模式等还很少见报道。本文研究了茂兰喀斯特森林林隙植被发育过程中的物种组成及种间关系的动态变化规律,为喀斯特森林群落结构动态及维持机制的研究提供理论基础。
1 研究区概况贵州省南部黔、桂交界处的茂兰国家级自然保护区(25°09′—25°20′ N,107°52′—108°05′ E),该区保存着世界同纬度地区仅有的原生性喀斯特森林。区内为典型的喀斯特峰丛地貌,最高海拔1 078.6 m,最低430 m,平均在800 m以上。成土母岩以中下石炭纪白云岩及石灰岩为主。当地年均气温15.3 ℃,7月均温26.4 ℃,1月均温8.3 ℃,≥10 ℃积温5 727.9 ℃,年均降雨量1 320.5 mm。全年平均相对湿度83%。土壤以黑色石灰土为主,土层浅薄,地面岩石裸露,土壤pH值7.5~8.0,有机质和全氮含量丰富(龙翠玲等,2005a)。
研究样地设在茂兰喀斯特森林自然保护区核心区原生性顶极常绿落叶阔叶林内,样地海拔750 m。以耐旱、喜钙的植被类型为主,林分郁闭度0.95。林冠层植被高10~20 m。该群落结构复杂,树种繁多,主要树种有圆果化香(Platycarya longipes)、翅荚香槐(Cladrastis platycarpa)、青冈(Cyclobalanopsis glauca)、椤木石楠(Photinia davidsoniae)、狭叶润楠(Machilus rehderi)等。下木及草本植物种类繁多。
2 研究方法 2.1 野外调查在茂兰保护区核心区选取典型地段设置标准地,面积约3.0 hm2。在样地内寻找林隙,辨认林隙形成木的种类,测量其胸径和高度。受条件限制,用倒木或枯立木腐烂级与附近林区1974年择伐时遗留的伐倒木腐烂程度进行对照,大致估测林隙形成木的年龄。对每棵边缘木做树冠投影图,冠空隙的面积大小采用坐标方格求算,扩展林隙大小由周围边缘木组成的多边形求算。分别记载每一林隙的乔木、灌木的种类、个体数、高度、胸径或基径和冠幅。在林隙中随机设置5个1 m×1 m的小样方调查草本、乔灌幼苗的种类、个体数和盖度等内容,共调查到林隙60个。在标准地中随机选设非林隙林分,设10 m×10 m的调查样地共20个,调查样地中1.5 m以上树木的种名、高度和胸径及所有苗木的种名、高度和基径,并在样地中随机设置5个1 m×1 m的小样方调查草本的种类和盖度。此外,规定胸径≤1 cm的树木为幼苗,1~10 cm者为幼树,>10 cm者为大树。
2.2 分析方法选取面积为100~200 m2的林隙35个,以10年为1年龄级,按10、20、30、40年(每个龄级的林隙数分别为8、10、9和8个)及非林隙5个林隙发育阶段分别统计物种的重要值Ⅳ及林隙阶段间物种的相似性系数Cj:Ⅳ=(相对密度+相对频度+相对显著度)/300 (乔木和灌木),Ⅳ=(相对密度+相对高度+相对盖度)/300 (草本和藤本);Cj=c/(a+b-c),c为2个群落共有的物种数,a为群落A中物种数,b为群落B中的物种数。
3 结果与分析 3.1 林隙植被发育过程中群落物种组成及优势度的变化表 1列出了林隙发育各阶段优势度较大的物种(表中物种重要值为各年龄阶段所有林隙的物种数统计所得)。采用目前应用最广泛的Raunkiaer生活型分类系统,确定各个种的生活型,把林隙植物分成乔木层、灌木层、草本层和藤本4个层次进行统计。从表 1可以看出,林隙物种的优势度随林隙年龄的增大有起伏,不同物种在不同林隙阶段成为优势种,且大部分物种的优势度在不同的林隙发育阶段上连续性较强,在发育时间为10、20、30和40年的序列上,林隙中乔木层优势种包括小叶柿(Diospyros dumetorum)、黄连木(Pistacia chinensis)、掌叶木(Handeliodendron bodinieri)和云贵鹅耳枥(Carpinus pubescens);灌木层优势种包括贵州悬竹(Ampelocalamus calcareus)、革叶铁榄(Sinosideroxylon wightianum)、湖北十大功劳(Mahonia confusa)和长梗核果木(Drypetes longipes);草本层优势种包括石韦(Pyrrosia lingua)、瓦韦(Lepisorus)、翠云草(Selaginella uncinata)和庐山楼梯草(Elatostema stewardii);藤本优势度最大的物种分别是鞘柄菝葜(Smilax stans)、四川清风藤(Sabia schumanniana)、藤黄檀(Dabergia hancei)和圆锥清风藤(Sabia paniculata)。在非林隙林分中,多数常绿乔木和灌木树种如青冈、川桂(Cinnamomum wilsonii)、革叶铁榄等的优势度比林隙后期阶段有明显增大的趋势,多数落叶乔木和灌木树种如掌叶木、朴树(Celtis sinensis)等的优势度随林隙年龄的增大而减小;石韦和藤黄檀等阳性耐旱的草本和藤本植物优势度随林隙年龄的增大而减小,而西南冷水花(Pilea plataniflora)和三叶崖爬藤(Tetrastigma hemsleyanum)等耐荫喜湿的草本和藤本植物优势度增加。贵州悬竹大量出现在林隙发育的早期阶段,随林隙年龄的增长,逐渐被其他物种取代,因此在郁闭林中没有其个体存在。由表 1还可知,乔木层的一些树种,如青冈、云贵鹅耳枥、小叶柿等,在林隙形成第10年时优势度均较高,第20年时达最大值,第30年时减小,至第40年时又增大,到林隙郁闭时在群落中占有一定的优势地位。而另一些树种,如掌叶木、圆果化香等在林隙形成初期优势度较小,第30年时达最大,在郁闭林分中优势度也较小。一些树种如角叶槭(Acer sycopseoides)、贵州泡花树(Meliosma henryi)等在林隙的发育过程中优势度变化和缓。另外一些灌木和小乔木,如南天竹(Nandina domestica)、羊蹄甲(Bauhinia variegata)等的优势度随林隙的发育缓慢增加,但在郁闭林林冠层没有其个体存在。除此之外,少部分乔木和灌木树种,如翅荚香槐、羊蹄甲等,在林隙的发育过程中优势度有缺失现象,茂兰喀斯特森林林隙树种的这种动态变化与各树种更新行为的差异是密不可分的。
|
|
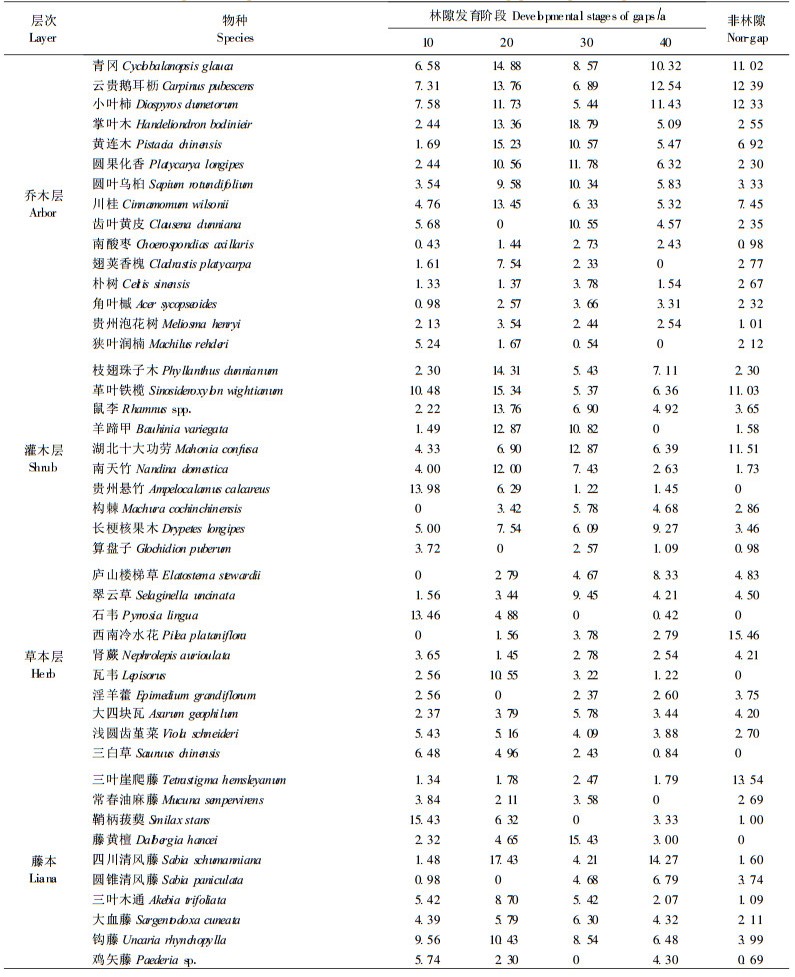
不同林隙阶段群落的共有种及相似系数的计算结果见表 2。由表 2可知,乔木层在林隙形成的10年内物种数最少,与其余各阶段的相似系数也很低,平均仅为0.24。林隙发育到第20年时,乔木层物种与其他林隙阶段的相似系数有所提高,平均达到0.28,说明这一阶段出现的乔木物种对林隙环境的适应能力有所加强。第30年时林隙内不仅乔木的物种数多,而且与其余各阶段的相似系数也较高,达0.40。第40年时林隙的乔木物种数有所下降,与其他林隙阶段的平均相似系数为0.38,表明林隙发育至40年时,林隙更新过程中出现的这些乔木树种在维持喀斯特森林乔木物种的连续性和稳定性方面具有一定的作用,其中一些种可能属于喀斯特森林中的优势乔木物种。到非林隙阶段时,只有2种仍保留在群落中,另外3种都已在林隙的更新过程中消失。
|
|
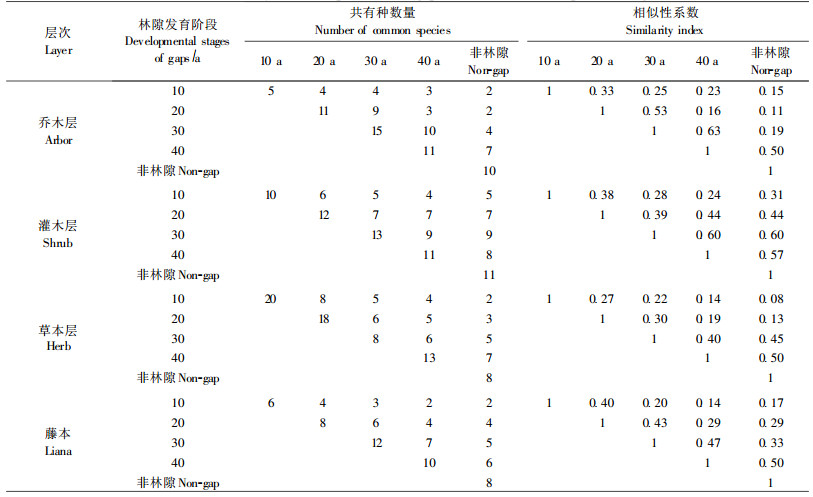
各林隙阶段灌木层物种的平均相似系数分别为0.30、0.41、0.47、0.46和0.48,其中20年林隙阶段多数灌木种类能够持续到非林隙阶段而不消失,说明这些物种对林隙环境适应能力较强,是林隙发育过程中的优势灌木物种。
草本植物种类在林隙发育过程中的持续性相对较差。在林隙初期(10年内)尽管草本数量多达20种,但当林隙发育至第20年时仅剩下8种,其余12种已经消失,并且只有2种持续到了非林隙阶段。由此可见,草本植物在恢复过程中物种更新十分频繁,早期的喜光草本物种很难适应郁闭后的荫蔽环境。20年林隙阶段的草本物种尽管数量也不少,有18种,但在林隙更新过程中的变化趋势与林隙初期阶段的草本物种十分相似。30年林隙阶段草本植物种类急剧下降,只有8种,但却还有5种能够持续到非林隙阶段,说明从30年林隙开始草本植物已经完成了对林隙环境的适应过程,大量的物种已经被淘汰,留存的草本以耐荫种类为主,对林隙后期阶段的荫蔽环境适应性较强。
藤本植物在林隙第10年时只有6种,且只有2种能够延续到非林隙阶段,其余4种已经消失,与其他林隙阶段的相似系数也较低,平均为0.23。林隙在第20年时种类有增加趋势,在第30年时增加到12种,有5种能到达非林隙阶段,与其他阶段的相似系数也较高,平均为0.36。在第40年时林隙与非林隙的物种组成类同,与其他阶段相似系数为0.35,表明林隙发育至第40年时,藤本植物种数也不再增加,优势种逐步确立。
3.3 林隙发育过程中物种的更新模式图 1为林隙中主要树种林冠层及其林下幼苗、幼树相对密度的变化趋势,从中可知其对林隙的填充过程,也体现了不同类型树种在林隙中更新行为的差异。每一树种代表一种林隙更新模式:1)以青冈为代表的树种,林隙形成初期就已存在大量的幼树和幼苗,在林隙发育的前30年幼树迅速增长,40年时多数幼树个体能够到达林冠层,并成为林冠层的主体,表明这些树种在郁闭林下形成幼苗,林隙形成后通过自我更新实现其更新,在林隙发育过程中有完整的种群结构,如川桂、小叶柿、狭叶润楠等树种属于此类型,多数为耐荫性较强的常绿树种;2)以黄连木为代表的树种,在林隙形成初期幼树不多见,但幼苗较多,当林隙发育到30年时,幼树最为丰富,幼苗逐渐减少,最终只有少数幼树个体能到达林冠层,且在郁闭林中无更新幼体存在,表明这些树种在林隙形成后产生大量幼苗,随林隙的发育通过自我更新和相互更新两种途径实现更新,属典型的林隙更新树种,少数种类在林隙发育过程中有完整的种群结构,如圆果化香、圆叶乌桕、云贵鹅耳枥、榆(Ulmus)、朴(Celtis)、榉树(Zalkova)等属于此类型,大部分是耐荫性较弱的落叶树种;3)在林隙形成的前20年,角叶槭等树种的幼苗、幼树稳定增长,30年后稍有下降,但仍保持一定幼苗、幼树数量,最终只有极少数个体到达林冠层,表明这些树种对林隙环境的变化不敏感,在林隙中形成幼苗,随林隙的发育主要通过相互更新模式进入林冠层,随林隙的发育少数种类具有完整的种群结构。贵州泡花树、南酸枣、球核荚NFDA3 (Viburnum propinquum)等属此类型,这些树种在群落中优势地位不显著,多数为中层小乔木和群落伴生种;4)湖北十大功劳等灌木树种的幼苗数量随林隙的发育而减少,幼树数量则缓慢增加,在林冠层中无个体存在。表明这些树种对林隙环境的变化反应虽活跃,但不能到达主林层,为林下层树种,随林隙的发育无完整的种群结构,如南天竹、算盘子、长梗核果木等灌木和少数亚冠层树种属此类型。

|
图 1 不同更新模式代表树种的相对密度 Fig. 1 Relative density of typical tree species in different generation categories |
茂兰喀斯特森林林隙发育过程中物种组成的动态变化格局反映了林隙植被发育过程中群落组成和生物多样性对这种变化的响应过程。林隙发育过程中由于生态环境不断变化,不同性质的物种对此作出不同的响应,表现为不同林隙阶段有不同的种类组成、数量特征和行为表现。在林隙形成初期,由于有较优越的光照条件和较小的竞争环境,林隙中幼苗、幼树数量较多,主要为一些喜光先锋树种,幼苗发生率高,死亡率也相对较高。Yamamoto等(1999)把树种在林隙中的更新方式分为自我更新和相互更新2种模式,自我更新指物种由同种个体更新的过程,而相互更新为一物种被另一物种更新的过程。多数阳性落叶树种在林隙形成初期虽然幼苗数量较多,但被其他一些树种取代的机率也较高,即死亡率高,最终只有少数个体能到达林冠层,且多数种类种群结构不完整,因此,多数落叶树种在林隙中采用相互更新模式实现其更新。常绿树种在林下形成幼苗,且耐荫性较强,待林隙形成后迅速生长,成为群落的优势种。因这类物种的竞争能力较强,在林隙的发育过程中很少会被其他种类所更新,常具有完整的种群结构,因此,常绿树种主要以自我更新方式实现其更新。在林隙的发育与填充过程中,自我更新和相互更新兼而有之,林隙的形成为不同更新方式和途径的物种的更新提供了场所。有研究表明(龙翠玲等,2005b):林隙更新过程中物种多样性的变化主要体现在一些阳性落叶树种的相互更新过程中,林隙形成初期,光照条件改善,土壤种子库和外来侵入的喜阳树种种子大量萌发,体现为林隙形成后物种多样性增加,随着林隙的发育,林分郁闭度增加,已不适合于耐荫性较小物种的生存,喜阳树种将被耐荫性较大的物种所更新,因而物种多样性降低。林隙的发生、发展和成熟,为不同生物学、生态学特性的物种提供了“中采式”竞争(lottery competition)的机会,使个体数在不同物种之间分布比较均匀,生态优势度较小,因而多样性指数较高,这是林隙干扰对物种多样性共存机制的作用所在。
在林隙的发育过程中,群落不同层次物种组成动态存在较大差异。经计算,在林隙发育过程中乔木层、灌木层、草本层和藤本植物的平均相似系数分别为0.31、0.42、0.27和0.32。可见,林隙发育过程中群落各层次的相似系数由大到小的顺序为:灌木>藤本>乔木>草本,灌木种类随林隙的发育具有较高的连续性和稳定性;草本植物则随着林隙环境条件的改变而出现较大的消亡和更新,显示出较大的波动性;乔木和藤本植物的稳定性和连续性介于灌木和草本植物之间。表 2还表明:每一林隙阶段总是与其下一阶段最邻近的群落具有最高的相似度,随着林隙的发育,群落之间的时间间隔越长,则相似系数越低,表明群落物种组成差异不断增加,群落之间的生态距离变远。群落相似系数的这种变化清楚地表明了林隙发育过程中物种组成结构的递进性和渐变性。对于以上的结果试作如下解释:由于受喀斯特森林小生境分布的不均匀性、不连续性的影响,植物个体和种群的分布有极大的随机性,体现了喀斯特生境基质对其上物种分布格局的深刻影响。乔木层树种由于个体植株高大,对生境营养空间要求较高,而喀斯特生境营养空间的有限性和不连续性却成为其分布的限制性因子,因而乔木在林隙发育过程中连续性较差。相对而言,灌木处于更新层,个体较小,但数量较多,对林隙环境变化的反应又十分活跃,因此灌木种类具有较高的连续性和稳定性。草本植物虽然种子扩散能力强,能较好地利用环境资源,但这种能力也增加了其分布的随机性,加上生活史周期短,因而在林隙发育过程中连续性最差,显示出较大的波动性。藤本植物由于具有较强的分蘖萌生能力和延伸性,连续性高于草本。
对不同年龄林隙的物种种间关系的研究,有助于对喀斯特森林物种共存机制的认识。研究结果表明:林隙的形成对茂兰喀斯特森林不同性质物种的分布、区系组成多样性和群落层次结构的维持有着重要影响,特别是林隙的时空尺度对喀斯特森林物种的分布起到筛选和分割作用,体现为不同性质的物种分布在不同年龄林隙中;除此之外,喀斯特地质地貌和生境特征也对物种的分布产生深刻影响。对于喀斯特森林这样一类特殊的生态系统而言,物种共存机制极其复杂,是生物与环境协同进化以及不同时空尺度的自然干扰综合作用的结果。因此,茂兰喀斯特森林植被具有其独特性,植物个体的生理生态特性、植物种间和种内关系的变化等方面的内容还有待于进一步的研究。
何永涛, 李贵才, 曹敏, 等. 2003. 哀牢山中山湿性常绿阔叶林林窗更新研究. 应用生态学报, 14(9): 1399-1404. DOI:10.3321/j.issn:1001-9332.2003.09.001 |
刘济明. 2000. 茂兰喀斯特森林主要树种的繁殖更新对策. 林业科学, 36(5): 114-122. DOI:10.3321/j.issn:1001-7488.2000.05.019 |
龙翠玲, 余世孝, 魏鲁明, 等. 2005a. 茂兰喀斯特森林干扰状况与林隙特征. 林业科学, 41(4): 13-19. |
龙翠玲, 余世孝, 熊志斌, 等. 2005b. 茂兰喀斯特森林林隙的植物多样性与更新. 生物多样性, 13(1): 43-50. |
王周平, 李旭光, 石胜友, 等. 2001. 重庆缙云山针阔混交林林隙树木更替规律研究. 植物生态学报, 25(4): 399-404. DOI:10.3321/j.issn:1005-264X.2001.04.003 |
熊小刚, 熊高明, 谢宗强. 2002. 神农架地区常绿落叶阔叶混交林树种更新研究. 生态学报, 22(11): 2001-2005. DOI:10.3321/j.issn:1000-0933.2002.11.029 |
臧润国, 余世孝, 刘静艳, 等. 1999. 海南霸王岭热带山地雨林林隙更新规律的研究. 生态学报, 19(2): 151-158. |
朱守谦. 1993. 喀斯特森林生态研究(Ⅰ). 贵阳: 贵州科技出版社, 1-11.
|
Arnaga L. 1988. Gap dynamics of a tropical cloud forest in northeastern Mexico. Biotropica, 20(3): 178-184. DOI:10.2307/2388232 |
Lertzman K P. 1992. Pattern of gap-phase replacement in subalpine, old-growth forest. Ecology, 73: 657-669. DOI:10.2307/1940772 |
Whitemore T C. 1989. Canopy gaps and the two major groups of forest trees. Ecology, 70: 536-538. DOI:10.2307/1940195 |
Yamamoto S, Nishimura N. 1999. Canopy gap formation and replacement pattern of major tree species among developmental stages of beech (Fagus crenata) stands, Japan. Plant Ecology, 140: 167-176. DOI:10.1023/A:1009713002039 |