 2007, Vol. 43
2007, Vol. 43文章信息
- 韩崇选, 张刚龙, 王明春, 张宏利, 杨清娥, 杨学军.
- Han Chongxuan, Zhang Ganglong, Wang Mingchun, Zhang Hongli, Yang Qing'e, Yang Xuejun.
- 黄土高原次生林改造林地鼢鼠与林木关系动态分析
- Dynamics Analysis of the Relationship between Zokor and Secondary Forests on the Loess Plateau
- 林业科学, 2007, 43(8): 71-79.
- Scientia Silvae Sinicae, 2007, 43(8): 71-79.
-
文章历史
- 收稿日期:2005-12-13
-
作者相关文章


自然事件的影响和人类活动的干扰,越来越多的大片森林破碎成彼此孤立的“森林岛屿”(Saunders, 1991; Donovan, 1997)。这种破碎化模式不仅形成了黄土高原今天较大面积的天然次生林,而且增加了植被的脆弱性(韩崇选等,2005a;2004b)。由于次生林结构类型单调,缺乏良好的经营管理,致使林业资源和经济建设陷入危困中(李国猷,1992;包晓斌,1994)。为了缓解这种状况,各地都进行了不同程度的次生林改造,以改变该区域森林资源现状(中国林学会,1984;张其保等,1984;佘济云等,2002)。但随着次生林改造的大面积进行和天然林保护工程的全面实施,林地鼢鼠危害问题逐渐突显,成为了影响次生林改造和天然林保护工程实施的主要因素(韩崇选等,1994a;2005a;2004b)。有关鼢鼠的研究报道很多(禹瀚,1956;郑宝赉等,1963;王祖望等,1975;郑生武,1980;王廷正等,1993;张堰铭,1999;王明春等,1997;2005;樊乃昌等,1990;江廷安,1998;韩崇选等,1994b;1995;2002;2005b), 奠定了鼢鼠可持续控制研究的基础。但是,由于鼢鼠发生规律的独特性和多样性,加之次生林生态系统结构的特殊性和各种物理、生物参数的复杂性,次生林改造后林地鼢鼠发生规律研究报道很少。笔者通过对次生林改造林地甘肃鼢鼠(Myospalax comsus)与林木的关系研究,评价不同次改模式的效果,为制定次生林改造和天然林保护工程的实施方案提供科学依据。
1 研究基地概况研究基地选择在陕西省宝鸡市麟游县的杨家沟和延安市的桥山、桥北林业局,属国家退耕还林工程和天然林保护工程的重点实施区域。杨家沟位于渭北高原中部,海拔1 350~1 520 m,全年日照平均时数为2 190.3 h,年平均气温为9.2 ℃,极端最高达37.5 ℃,极端最低为-55.1 ℃,无霜期平均178 d,年平均降雨量640.4 mm,年蒸发量为806.0 mm。区内农林交错分布,牛羊啃食比较严重,为甘肃鼢鼠的重发区;土壤为垆土和森林黄土性黄土。植被破坏严重,仅残存有零星的次生乔、灌木分布,森林覆盖率为19.0%。桥山和桥北林业局位于陕北高原南部,年均气温8.6 ℃,年均降雨量561.3 mm,土壤主要是黑垆土和黄绵土,次生林以山杨(Populus davidiana)、白桦(Betula platyphylla)、辽东栎(Quercus liaotungensis)为主。林地与灌木疏林、草地、农田多呈镶嵌分布。造林树种以油松(Pinus tabulaeformis)(ch)、侧柏(Platycladus orientalis)(ca)、刺槐(Robinia pseudoacacia)(sc)和沙棘(Hippophae rhamnoides)为主,其次是少量的山桃(Prunus davidiana)、山杏(Armeniaca vulgaris var. ansu)、仁用杏(Armeniaca vulgaris×A. sibirica)和苹果(Malus pumila)等(韩崇选等,2003a; 2003b;2004a;2004c)。
2 材料与方法1992—1997年,结合中德合作陕西造林工程项目和油松良种基地建设,在项目区采取5种模式进行次生林改造试验(韩崇选等,1994a;王明春等,2004;杨学军等,2001)。造林树种为油松、侧柏和刺槐。1)全面清坡造林,选择地势平坦或坡度不大、土层深厚且比较肥沃的次生林,将林木全部清除,采取水平阶整地重新造林(over-all felling, of)。2)带状清坡造林分为2种形式,一种为水平带状清坡造林(level felling, lf),按等高线每隔4 m宽的次生林带,砍除2 m宽地带上的所有林木,在伐带上按2.0 m的株距造林;同时对保留带上的次生林木进行疏伐,按合理密度保留杂灌;一种为竖向清坡造林(vertical felling, vf),清坡宽度为20 m,保留带宽度为5 m,然后在清坡带上水平阶整地造林。3)块状清坡造林(patch felling,pf),在山坡的中部,每隔5 m,设置30 m×30 m样方,清除样方中的所有林木,采用等高线水平阶整地造林,株行距2 m×3 m。4)林下更新造林(regeneration, ra),块状清除后挖穴植苗,清除面积4 m2,补苗750~1 200株·hm-2,以后3年,每年对新植幼树进行块状抚育1次。造林后每年10月,采用切封洞法调查每块样地的封洞数,同时在对照林地采用大面积捕尽法确定洞口系数和种群结构。调查各样地主要造林树种的被害率、被害死亡率。对调查的数据分类整理,采用系统分析法,比较次生林改造后林地鼢鼠发生规律与次生林鼢鼠发生规律的异同,评价5种次生林改造模式对林地鼢鼠抑制的效果。
3 结果与分析 3.1 不同改造模式林木被害程度分析不同改造模式,林木被害死亡率不同,同一改造模式,不同树种的不同定植年限,林木被害死亡率也有差异(表 1)。
|
|
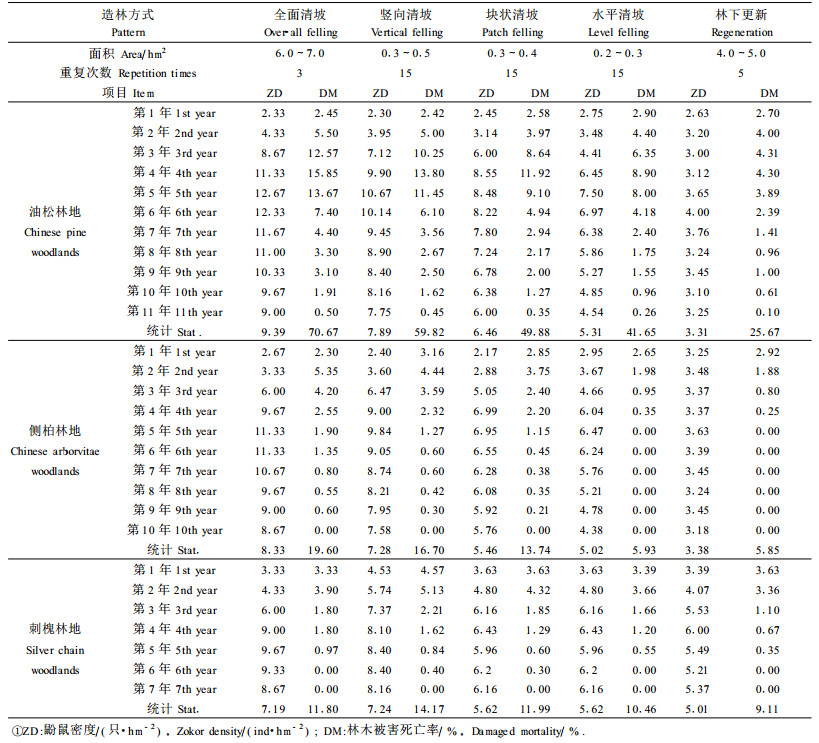
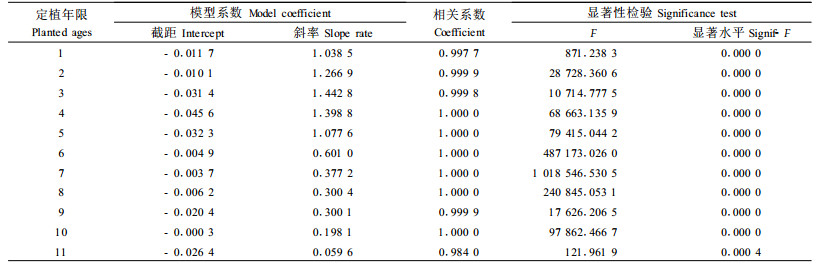
对表 1中的林木被害死亡各年累加值按时间序列分析。5种改造模式造林后,油松的逐年累计被害死亡率均遵从Logistic模型变化规律,其模型如下:
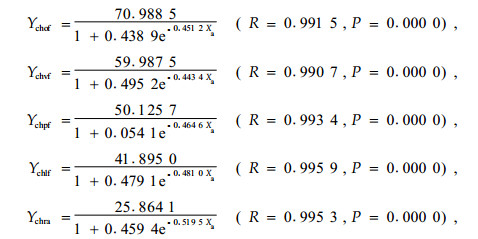
|
从模型分析,改造初期,林木被害死亡率随年限变化的速率较小;随着定植年限的增加,死亡率的增加速率逐渐增大,当累计死亡率达到其稳定值(最大值)一半时,也就是定植后的第4年,其增加速率达最大;当累计死亡率超过其稳定值的一半时,死亡率随定植年限增加的速度逐步降低,并逐渐接近于零。虽然林木被害死亡率的变化规律相同,但不同改造模式下林木的被害程度差异很大。模型预测5种模式油松的最大累计被害死亡率依次为70.99%、50.13%、50.13%、41.90%和25.86%,与实际值70.67%、59.82%、49.88%、41.65%和25.67%十分接近。对不同模式林木被害程度进行ANOVA分析和LSD配对比较分析可知,全面清坡模式与竖向清坡模式差异不显著(P=0.390),与块状清坡模式有一定的差异(P=0.102),与水平带状模式差异显著(P=0.025),与林下更新模式差异极显著(P=0.001);竖向清坡模式与块状清坡模式差异不大(P=0.430),与水平带状模式差异不显著(P=0.156),与林下更新模式差异极显著(P=0.009);块状清坡模式与水平带状模式无差异(P=0.523),而与林下更新模式有差异(P=0.059);水平带状模式与林下更新模式差异不显著(P=0.203)。鼢鼠对定植11年以下的油松均可能造成危害,定植2~6年的林木被害程度较大,5种模式林木在该时间段内被害死亡率占总死亡率的比例依次为77.81%、77.90%、77.32%、76.42%和73.59%。
3.1.2 侧柏的被害程度进程分析改造后侧柏的连年累计被害死亡率变化规律与油松的基本相同。全面清坡林木被害程度与竖向清坡和块状清坡模式差异不显著(P=0.443),与水平带状和林下更新模式差异极显著(P=0.000);竖向清坡模式与块状清坡模式无差异(P=1.000),竖向清坡和块状清模式与水平带状和林下更新模式差异极显著(P=0.002);水平带状与林下更新模式无差异(P=0.983)。相对全面清坡造林模式,后4种模式对鼢鼠的拟制效果分别为14.80%、29.90%、69.74%和70.15%。对林木的被害程度进程进行模型分析,全面清坡、竖向清坡和块状清坡造林模式,林木累计被害死亡率既遵从S-模型,又符合Logistic模型;水平带状和林下更新模式造林符合Inverse模型,也符合Logistic模型,其模型如下:
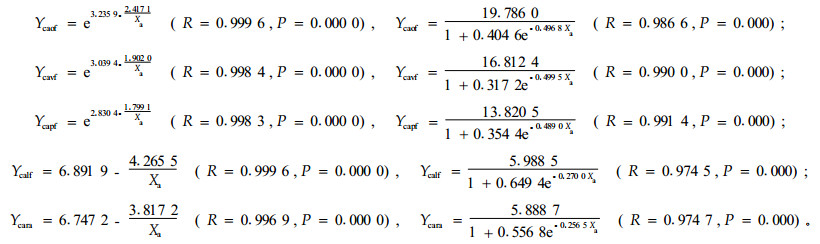
|
从模型回测分析,S-模型与全面清坡造林后1~5年的林木累计死亡率相关较紧密,其模型的有效最小值为2.31%,与实际值2.30%几乎相等,而模型的有效极大值和极大值分别为20.37%和25.43%,与侧柏的最大连年累计死亡率1.60%相差显著;Logistic模型对5年以后的林木被害死亡率预测准确,其模型的有效极大值和极大值为19.64%和19.79%,与侧柏的最大连年累计死亡率19.60%相差不显著,而模型的有效极小值为3.98%,与侧柏第1年的被害死亡率差异显著。竖向和块状清坡造林模式2模型的回测分析结果与全面清坡造林的相似。Inverse模型对水平带状和林下更新模式第1年的回测值为2.63%和2.93%,与实际值2.65%和2.92%十分接近,而有效极大值和极大值分别为5.83%和6.89%与5.79%和6.74%,与侧柏第4年的累计死亡率5.93%和5.85%差异显著;Logistic模型的有效极小值分别2.92%和3.20%,与实际值差异较大,而有效极大值和极大值依次为5.87%、5.99%和5.81%、5.89 %,与实际值5.93%和5.85%差异不显著。
3.1.3 刺槐的被害程度分析刺槐被害累计死亡率随定植年限的变化,前4种造林模式可用S-模型和Logistic模型描述,林下更新模式可用Inverse模型和Logistic模型表示,其模型如下:
|
模型回测分析结果与侧柏的一致。对5种模式的林木被害程度进行ANOVA分析和LSD配对比较分析,仅竖向清坡与更新模式差异显著(P=0.045),与水平带状模式有一定的差异(P=0.099)。不同模式刺槐被害程度与油松和侧柏有所不同,表现为竖向清坡造林模式最重,其次是块状清坡、全面清坡和水平带状清坡模式,林下更新模式刺槐被害最轻。造成变化的原因主要是造林初期林地鼠口密度的差异。消除造林初期林地鼠口密度差异的影响,竖向清坡模式定植6年的刺槐平均被害累计死亡率为11.32%,块状清坡、水平带状和林下更新模式依次为10.99%、10.27%和8.95%。
3.2 林木被害程度模型 3.2.1 油松被害程度模型不同改造模式,单个鼢鼠对油松危害死亡率的变化规律基本一致(图 1-1)。ANOVA分析和LSD配对比较分析结果证明,单个鼢鼠在同年各模式下对林木的危害程度是一致的(F=0.000,P=1.000),但年度间差异极显著(F=6 532.926,P=0.000)(图 1-1)。同一年林木被害死亡率与林地鼠口密度呈极显著的正相关(表 2),模型截距近似于0,斜率随定植年限遵从Cubic-模型(Y=0.612 6+0.592 7X-0.137 0X2+0.007 2X3,R=0.970 5,F=37.764 4,P=0.000 1)变化规律。从直线回归模型斜率分析,单个鼢鼠对油松的危害程度造林后1~3年有一个上升过程,第3年以后,危害强度逐年下降。单个鼢鼠对油松的危害程度主要取决于林木定植年限,而与改造模式和鼢鼠密度关系不明显。

|
图 1 油松被害程度模型分析 Fig. 1 The mode analysis chart of damaged degree of Chinese pine 模式中,1:全面清坡;2:竖向清坡;3:块状清坡;4:水平带状;5:林下更新。In the modes, 1: Over-all felling; 2: Vertical felling; 3: Patch felling; 4: Level felling; 5: Regeneration.下同。The same below. |
|
|
对不同模式林地鼢鼠密度进行ANOVA分析和LSD配对比较分析,各模式林地鼢鼠密度存在极显著的差异(F=14.905,F=0.000),其中,全面清坡与竖向清坡模式有一定差异(P=0.088),但差异不显著,而与其他3种模式差异极显著(P<0.01);竖向清坡与块状清坡不显著(P=0.095),而与水平带状和林下更新模式差异极显著(P<0.01);竖向清坡与水平带状模式不显著(P=0.175),而与林下更新模式差异极显著(P=0.001);水平带状与林下更新差异显著(P=0.028)。不仅不同模式间有差异,不同年份间林地鼢鼠密度也存在差异(F=2.539,F=0.013),定植当年的林地密度除与第2年差异不显著外(P=0.492),与以后各年均存在着显著的差异(P=0.000~0.045),第2年与4~8年差异显著(P=0.004~0.031),而与其他各年差异不显著(P<0.050);第3年与以后各年差异不显著(P=0.098~0.879)。不同模式林木被害程度的ANOVA分析和LSD配对比较分析结果与林地鼢鼠密度结果一致,也表现为各模式间(F=3.774,F=0.009)和各年度间(F=9.655,F=0.000)均存在极显著差异(图 1-2,图 1-3)。对林木被害死亡率(Ydm)与定植年限(Xpa)、改造模式(Xsm)、鼢鼠密度(Xzd)和表 2中模型斜率(Xs)进行多元逐步回归分析(图 1-2,图 1-3)。其模型如下:
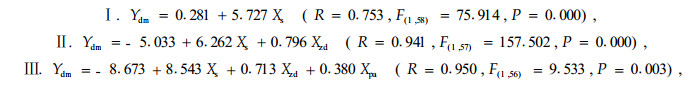
|
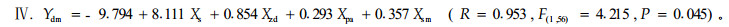
|
模型参数显著性检验证明:模型Ⅰ的常数项相关不显著(t=0.505,P=0.616),Xs项相关极显著(t=8.713,P=0.000);模型Ⅱ和模型Ⅲ各参数均极相关(t值依次为-9.812、18.180、12.550、-6.823、10.612、3.091,对应的P值除模型Ⅲ的Xpa项为0.003外,其余均为0.000);模型Ⅳ的前3项参数极相关(t=-7.248、10.004、9.502,P=0.000),后2项显著相关(t=2.312、2.053,P=0.025、0.045)。各参数的单相关系数依次为0.753、0.466、-0.568和-0.325。模型Ⅰ反映了油松被害死亡率与其各年单个鼢鼠危害强度的关系,其实质是表现了油松被害死亡率与林地鼢鼠密度和油松定植年限的关系。模型Ⅱ主要反映了林木被害死亡率与单个鼢鼠和其种群的关系。模型Ⅲ引入了定植年限变量,表现了鼢鼠个体和种群对油松综合作用的年度变化。模型Ⅳ引入改造模式变量,证明林木被害死亡率不仅取决于林地鼢鼠个体对其的危害强度和鼢鼠种群密度,也取决于油松的定植年限和改造模式。
3.2.2 侧柏被害程度模型单个鼢鼠对侧柏危害死亡率的变化规律,不同模式间有所差异(图 2-1)。对其数据进行ANOVA分析和LSD配对比较分析,定植1~3年不同模式间差异不显著(F=1.225,P=0.360),其中,全面清坡与竖向清坡和块状清坡模式相似度较大(P=0.947、0.939),而与水平带状和林下更新模式有一定差异(P=0.166、0.176);定植4年以后,不同模式间差异明显(F=3.259,P=0.025),其中,全面清坡与竖向清坡模式相似性较弱(P=0.613),与块状清坡模式相似性较强(P=0.856),而与水平带状和林下更新模式差异显著(P=0.016、0.017);竖向清坡与块状清坡模式有一定的相似性(P=0.745),而与水平带状和林下更新模式差异较大(P=0.050、0.054);块状清坡与水平带状和林下更新模式差异显著(P=0.024、0.025);水平带状与林下更新模式相似性显著(P=0.971)。不仅不同模式间,单个鼢鼠对侧柏的危害死亡率变化较大,而且不同定植年度间单个鼢鼠对侧柏的危害程度差异很大(F=24.123,P=0.000)。对单个鼢鼠对侧柏危害速率(Yv)与定植年限(Xpa)、改造模式(Xsm)和林地鼢鼠密度(Xzd)进行多元逐步回归分析,其模型如下:

|
图 2 侧柏被害程度模型分析 Fig. 2 The mode analysis chart of damaged degree of Chinese arborvitae |
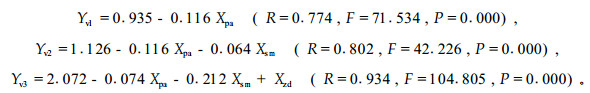
|
从模型可看出,对单个鼢鼠危害速率影响较大的林木定植年限首先进入模型,接着是改造模式参数进入模型,最后是鼢鼠密度参数进入模型。三者与危害速率呈负相关,其中,林木定植年限和鼢鼠密度与单个鼢鼠危害速率相关极显著(r=-0.774、-0.463,P=0.000),改造模式相关不显著(r=-0.210,P=0.072)。这说明影响单个鼢鼠对侧柏危害强度的首要因素是林木定植年限,其次是改造模式,最后是林地鼢鼠密度。这一结论与油松的有所差异。
林地鼢鼠密度和林木被害死亡率的ANOVA分析和LSD配对比较分析结果与油松的分析结果基本一致,但差异程度不同(图 2-2,图 2-3)。对于鼢鼠密度,各模式间差异极显著(F=9.1 21,P=0.000),其中,全面清坡与竖向清坡模式差异不显著(P=0.295),而与其他模式差异极显著(P=0.003、0.001、0.000);竖向清坡与后3种模式差异较大(P=0.048、0.015、0.000);块状清坡与水平带状模式差异不显著(P=0.621),而与林下更新模式差异显著(P=0.025);后2种模式差异不显著(P=0.075)。对于危害死亡率,不同模式间有一定差异(F=2.137,P=0.092),其中,第1种模式与竖向清坡和块状清坡模式差异不显著(P=0.634、0.339),而与后2种模式差异显著(P=0.029、0.028);竖向清坡、块状清坡和水平带状模式与其后模式差异不显著(P=0.080~0.990)。不同定植年限间,鼢鼠密度差异均显著(F=2.759,P=0.013),林木被害死亡率差异极显著(F=10.496,P=0.000)。对林木被害死亡率(Ydm)与定植年限(Xpa)、改造模式(Xsm)、鼢鼠密度(Xzd)和单个鼢鼠对林木的危害速率(Xv)进行多元逐步回归分析(图 2-2,图 2-3)。其模型如下:
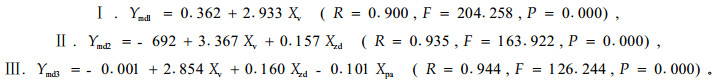
|
林木被害死亡率与单个鼢鼠危害速率呈显著的正相关(r=0.900,P=0.000),与鼢鼠密度呈不显著的负相关(r=-0.191,P=0.090),而与定植年限和改造模式呈极显著的负相关(r=-0.774、-0.386,P=0.000、0.003)。模型Ⅰ显示了林木被害死亡率与单个鼢鼠危害速率的关系,其实质是揭示了林木被害程度与林木定植年限、改造模式和鼢鼠密度的关系。模型Ⅱ反映了鼢鼠个体和种群密度对林木被害死亡率的影响。模型Ⅲ说明林木被害死亡率不仅取决于林地鼢鼠的综合作用,也取决于林木的定植年限。
3.2.3 刺槐被害程度模型单个鼢鼠对刺槐的危害程度与改造模式没有差异(F=0.009,P=0.999),而与定植年限和鼢鼠密度差异极显著(F=826.743、20.444,P=0.000)。其中定植1~5年与各年的两两差异均十分显著(P=0.000~0.005),定植6年和7年的差异不显著(P=0.207)。对单个鼢鼠的危害强度指标与林木定植年限、改造模式和鼢鼠密度指标进行模型分析,其适合模型如下:
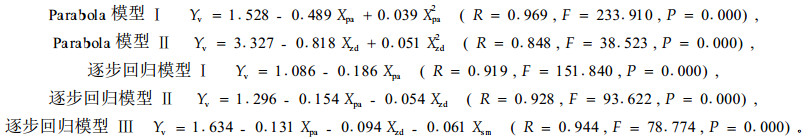
|
从Parabola模型分析,模型Ⅰ表述了定植年限与单个鼢鼠危害速率的关系,表现为速率随定植年限逐渐降低,且降低幅度逐年减小(图 3-1)。模型Ⅱ反映了鼢鼠密度与单个鼢鼠危害速率的关系,表现为随林地鼢鼠密度增加,单个鼢鼠危害速率有降低的趋势。从逐步回归模型分析,模型Ⅰ表示鼢鼠危害速率与林木定植年限呈极显著的负相关(r=-0.911,P=0.000)(图 3-1)。模型Ⅱ显示单个鼢鼠的危害速率随着林木的增长与鼢鼠密度呈极显著的负相关(r=-0.730,P=0.000)。模型Ⅲ反映了鼢鼠危害速率伴随林木的增长和林地鼢鼠密度的增加,与改造模式呈负相关,但相关不显著(r=-0.029,P=0.443)。从同年的数据分析,定植1~5年,林木被害死亡率与林地鼢鼠密度呈极显著的正相关(表 3)。其斜率与林木定植年限遵从Yv=1.487-0.465Xpa+0.036Xpa2的Pa rabola模型变化规律(R=0.974,F=277.712,P=0.000 0)。

|
图 3 刺槐被害程度模型分析 Fig. 3 The mode analysis chart of damaged degree of Silver chain |
|
|
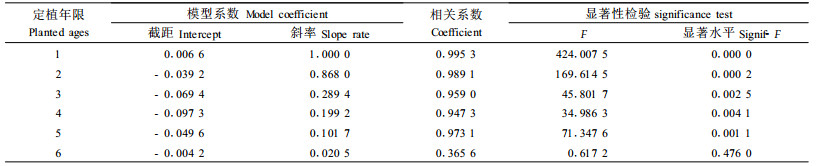
从林地鼢鼠密度和林木被害程度分析,不同改造模式,林地鼢鼠密度有一定差异(F=2.470,P=0.056),仅全面清坡和竖向清坡与林下更新模式差异显著(P=0.037、0.017)。林木被害程度不同模式间差异不显著(P=1.779、0.161),只有竖向清坡与林下更新模式差异显著(P=0.019)(图 3-2,图 3-3)。但两者年度间差异均十分显著(F=6.117、17.225,P=0.000)。对林木被害死亡率(Ydm)与定植年限(Xpa)、改造模式(Xsm)、鼢鼠密度(Xzd)、单个鼢鼠对林木的危害速率(Xv)表 3中模型斜率(Xs)进行多元逐步回归分析。其模型如下:
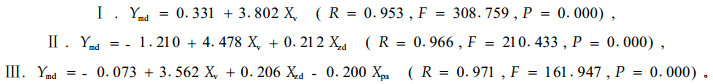
|
模型中,除模型Ⅲ的常数项不显著外(t=-0.109,P=0.914),其他参数均显著(P=0.000~0.029)。林木被害死亡率与单个鼢鼠危害速率和斜率呈极显著的正相关(r=0.953、0.949,P=0.000),与定植年限和鼢鼠密度为极显著的负相关(r=-0.913、-0.589,P=0.000)。
4 结论与讨论有害生物与目的植物的关系是有害生物优化管理决策和生态修复的主要依据和基础性工作,一直受到人们的关注(Donovan, 1997; 韩崇选等,2005a)。有关鼠类与林木关系的研究很多,但多数是建立在害鼠预测预报和经济阈值研究基础上的,公认的观点是害鼠对林木危害程度取决于林地鼠口密度、林木种类和定植年限等(韩崇选等,1994a; 2005b; 1995)。但忽视了害鼠个体与种群的相互作用和不同环境条件的影响,缺乏系统的研究证明。本研究发现:改造后,林木被害程度随定植年限的变化总体遵从Logistic模型,侧柏在全面清坡、竖向清坡和块状清坡模式下,也符合S-模型,在水平带状和林下更新模式下也符合Inverse模型,刺槐在前3种模式下,也符合S-模型,后2种模式下,也符合Inverse模型。对于油松,鼢鼠危害的年限为定植1~11年,其中2~6年被害最严重。全面清坡造林模式被害程度较高,林下更新造林模式被害程度较低。若以全面清坡造林模式为对照,其他4种模式对鼢鼠的预防效果分别为15.35%、29.42%、41.06%和63.68%。对于侧柏,前3种改造模式造林后9年内都可能受到鼢鼠的危害,其中造林后1~5年受害较重,造林第2年最为严重;而后2种模式较轻。对于刺槐,鼢鼠主要危害定植1~5年的刺槐,其中前4种模式定植2年的刺槐受害较重,林下更新模式第1年较重。与全面清坡清坡造林相比,对鼢鼠危害的预防效果分别为4.07%、6.86%、12.97%和24.15%。显然低于油松和侧柏相应的预防效果。
次生林改造后,鼢鼠对油松、侧柏和刺槐的危害程度差异很大。单个鼢鼠对油松和刺槐的危害程度主要取决于林木的定植年限,而与改造模式和鼢鼠密度无关。林木被害程度主要取决于林木定植年限和林地鼢鼠的种群密度,造林后1~3年危害程度有一个上升过程,第3年以后,危害强度逐年下降;对刺槐的危害程度随定植年限逐渐降低,但幅度逐年减小。同一年林木被害程度的差异,主要是由于不同改进模式林地鼢鼠种群密度的差异造成的,不同定植年限林木被害程度的差异主要是鼢鼠个体对林木危害强度差异和林地鼢鼠密度变化的结果。单个鼢鼠对侧柏的危害程度除取决于林木定植年限外,还取决于改造模式和林地鼢鼠密度。同一年林木被害死亡率与鼢鼠密度呈极显著的正相关,不同年限的与鼢鼠密度呈极显著的负相关。
林木被害程度的这种变化,反映了林木特性和次改幼林生物群落的演替关系,包括生物和非生物间关系的变化。次改后,原有的植被发生了变化,随着林木的生长,林下植被不断发生更替,林内光照、土壤湿度、气温等非生物因素也发生了很大变化,这些变化影响了林地鼢鼠的数量和分布,反过来又使林木被害程度发生变化。次生林改造初期,林下植被基本保持了原次生林草本植被的特点,主要是以白羊草(Bothriochloa ischaemum)为优势种的群落,鼢鼠喜食的阳性植物在林内呈块状核心分布,相对贫乏。鼢鼠密较低,分布相对集中,对幼树危害相对较轻,但由于幼树根系不发达,鼢鼠一旦觅食,一次可将根系全部吃光,只剩下树干,被害死亡率极高。随着林木的增长,林分郁闭度增加,林下鼢鼠喜食的萎陵菜(Potentilla chinensis)、二裂萎陵菜(P.bifurca)、苦荬菜(Ixeris denticulate)等逐渐扩散,覆盖度和组成比显著增加,鼢鼠密度逐渐上升,并从原分布地沿着水平阶整地带向周围林地扩散,聚集程度逐渐降低,取食林木的机会增大,被害率增高,但由于林木逐渐长大,鼢鼠一般一次不能啃食全部根系,形成部分林木虽然被害但未死亡,只是生长势减弱,其被害死亡率较幼树的低。随着林木的进一步增大,郁被度增加,林内光照减少,湿度增大,林下草本植被分布相对集中,有向团块状分布格局转化的趋势,出现以白羊草和长芒草(Stipa bungeana)为优势种的群落。鼢鼠喜食的萎陵菜、二裂萎陵菜、苦荬菜等植物的覆盖度和组成比逐渐降低。鼢鼠密度也逐年降低,聚集程度逐年增强,对林木的危害程度也逐渐降低。
包晓斌. 1994. 黄土高原土石山区天然次生林开发培育模式探讨. 中国水土保持, (2): 26-27. |
樊乃昌, 景增春, 周文扬. 1990. 高原鼢鼠的侵占行为及防治的新途径. 兽类学报, 10(2): 114-120. |
韩崇选, 胡忠朗, 陈孝达, 等. 1994a. 桥山林区甘肃鼢鼠发生规律研究. 陕西林业科技, 4: 23-29. |
韩崇选, 胡忠朗, 陈孝达, 等. 1994b. 甘肃鼢鼠对油松危害动态经济阈值研究及应用. 西北林学院学报, 9(3): 45-52. |
韩崇选, 胡忠朗, 杨学军, 等. 1995. 林地甘肃鼢鼠空间格局研究. 西北林学院学报, 10(1): 74-79. |
韩崇选, 王明春, 杨学军, 等. 2002. 安全型无公害灭鼠剂—克鼠星的研究. 西北林学院学报, 17(3): 44-47. DOI:10.3969/j.issn.1001-7461.2002.03.012 |
韩崇选, 杨学军, 王明春, 等. 2003a. 关中北部塬区林地啮齿动物群落多样性变化研究. 陕西师范大学学报:自然科学版, 31(专辑): 177-183. |
韩崇选, 杨学军, 王明春, 等. 2003b. 关中北部塬区林地啮齿动物群落结构和生态位研究. 陕西师范大学学报:自然科学版, 31(专辑): 184-190. |
韩崇选, 吕复扬, 卜书海, 等. 2004a. 陕西林区啮齿动物群落多样性研究. 西北林学院学报, 19(3): 99-104. |
韩崇选, 杨学军, 李金钢, 等. 2004b. 农林啮齿动物灾害的环境修复与安全诊断. 杨凌: 西北农林科技大学出版社, 160-380.
|
韩崇选, 杨学军, 王明春, 等. 2004c. 陕西林区草兔空间格局及区域变化研究. 西北农林科技大学学报:自然科学版, 32(11): 65-72. |
韩崇选, 李金钢, 杨学军, 等. 2005a. 中国农林啮齿动物综合管理. 杨凌: 西北农林科技大学出版社, 78-79; 197-234.
|
韩崇选, 杨学军, 王明春, 等. 2005b. 林区啮齿动物群落管理中的生态阈值研究. 西北林学院学报, 20(1): 156-161. |
江廷安. 1998. 陕北黄土高原啮齿动物的区系及鼢鼠的防治对策. 水土保持通报, 18(1): 48-53. DOI:10.3969/j.issn.1000-288X.1998.01.011 |
李国猷. 1992. 北方次生林经营. 北京: 中国林业出版社, 93-112.
|
佘济云, 曾思齐, 李志辉, 等. 2002. 湘东丘陵区马尾松低质低效次生林改造经营模式研究. 湖南林业科技, 29(1): 6-9. DOI:10.3969/j.issn.1003-5710.2002.01.002 |
王明春, 韩崇选, 胡忠朗, 等. 1997. 甘肃鼢鼠取食节律及对不同饵料喜食性的研究. 西北农业大学学报, 25(2): 37-41. DOI:10.3321/j.issn:1671-9387.1997.02.007 |
王明春, 韩崇选, 杨学军, 等. 2004. 林区甘肃鼢鼠危害特征及生态控制对策. 西北林学院学报, 19(3): 105-108. DOI:10.3969/j.issn.1001-7461.2004.03.030 |
王明春, 韩崇选, 宗德禄, 等. 2005. 无公害灭鼠剂克鼠星防治中华鼢鼠试验研究. 西北农林科技大学学报:自然科学版, 33(5): 39-43. |
王廷正, 李晓晨. 1993. 甘肃鼢鼠和中华鼢鼠繁殖特性的研究. 兽类学报, 13(2): 153-155. |
王祖望, 梁杰荣, 李俊荣, 等. 1975.鼢鼠数量与地面痕迹的关系//青海省生物研究所.灭鼠和鼠类生物学研究报告: 第2集.北京: 科学出版社, 83-93
|
杨学军, 韩崇选, 王明春, 等. 2001. 林业生态措施在鼠害控制中的应用. 西北林学院学报, 16(3): 76-79. DOI:10.3969/j.issn.1001-7461.2001.03.020 |
禹瀚. 1956. 陕北鼢鼠的初步研究. 西北农学院学报, (4): 59-65. |
张其保, 方亮. 1998. 大兴安岭岭南次生林改造方式与结构的研究. 内蒙古林学院学报:自然科学版, 20(3): 37-42. |
张堰铭. 1999. 捕杀对高原鼢鼠种群年龄结构及繁殖的影响. 兽类学报, 19(3): 204-211. DOI:10.3969/j.issn.1000-1050.1999.03.006 |
郑宝赉, 蔡桂全, 周乃武, 等. 1963. 滹沱河流域上游地区鼢鼠数量分布的调查研究. 动物学报, 18(1): 21-27. |
郑生武. 1980. 中华鼢鼠的繁殖研究. 动物学研究, 1(4): 465-477. |
中国林学会. 1984. 次生林经营技术. 北京: 中国林业出版社, 2-28.
|
Donovan T M, Jones P W, Annand E M, et al. 1997. Variation in local-scale edge effects:mechanisms and landscape context. Ecology, 78: 2064-2075. DOI:10.1890/0012-9658(1997)078[2064:VILSEE]2.0.CO;2 |
Saunders D A, Hobbs R J, Margules C R. 1991. Biological conservation of ecosystem fragmentation:a review. Conservation Biology, 5: 18-32. DOI:10.1111/j.1523-1739.1991.tb00384.x |