 2007, Vol. 43
2007, Vol. 43文章信息
- 韦小丽, 喻理飞, 朱守谦, 徐锡增.
- Wei Xiaoli, Yu Lifei, Zhu Shouqian, Xu Xizeng.
- 土壤干湿交替对青檀幼苗生理及生长的影响
- Effect of Soil Drying-Wetting Alternation on Physiology and Growth of Pteroceltis tatarinowii Seedlings
- 林业科学, 2007, 43(8): 23-28.
- Scientia Silvae Sinicae, 2007, 43(8): 23-28.
-
文章历史
- 收稿日期:2006-05-26
-
作者相关文章



2. 南京林业大学森林资源与环境学院 南京 210037
2. College of Forest Resources and Environment, Nanjing Forestry University Nanjing 210037
青檀(Pteroceltis tatarinowii)是国家三级保护植物,具有耐干旱瘠薄,喜钙质土壤的特性,是石灰岩山地造林的先锋树种。其木材坚硬致密而细致,是高级家具用材;檀皮是制作宣纸的高级原料,也是人造棉的主要原料之一;青檀叶营养丰富,是优质饲料(丁佐龙等,1997)。近年来,对于青檀的研究,涉及种子生理、育苗技术、人工林栽培管理和檀皮质量与加工(程孝霞等,2005;方升佐等,2001;2002; 洑香香等,2002)等方面,对青檀的水分生理、耐盐生理也作了一定的尝试(侯常英等,2003;Fang et al., 2006)。发育在湿润地区的喀斯特地貌是一种多变水环境,临时性水分胁迫频繁发生,但持续时间短,很容易为频繁的降雨所解除(朱守谦,2003)。因此,喀斯特森林树种总是在反复“干旱-复水”的环境中生存。本文以青檀幼苗为材料,采用盆栽试验人工模拟喀斯特地区常见的干湿交替环境,研究其在干湿交替环境下主要生理指标和生长指标的变化,以及幼苗在复水后的恢复能力,旨在阐明干湿交替环境对青檀幼苗生理指标及生长的影响,为喀斯特地区青檀人工栽培技术措施的制定提供理论依据。
1 材料与方法 1.1 试验材料试验材料为青檀当年生播种苗。育苗土壤是从石灰岩山地(贵阳花溪)挖取的白云质灰岩发育的土壤。原状土水分物理性质为:毛管持水量51%,田间持水量45.1%,密度1.007 g·cm-3,毛管孔隙度51%,非毛管孔隙度9%;土壤化学性质为:pH7.2,全氮0.169 g·kg-1,全磷0.112 g·kg-1,速效钾4.809 mg·kg-1,速效磷39.31 mg·kg-1,碱解氮0.303 1 g·kg-1,有机质含量1.532 g·kg-1。用高15 cm,口径14 cm塑料花盆装土,装土前对每个花盆编号称质量,各装入干土1 000 g(精度0.1 g)。于2004年3月8日播种,每盆播种3~5粒种子。出苗后通过间苗每盆保留2株生长一致的幼苗,于塑料大棚中培养,处理前按常规管理。
1.2 试验设计试验分别在幼苗生长初期(6月25日—8月4日)和速生期(8月9日—9月9日)进行(生长期是根据幼苗在当地的生长情况划分),每个生长时期选苗高、地径相对一致的幼苗进行处理,其中生长初期处理幼苗平均苗高、地径分别为(15.4±0.29) cm、(0.101±0.007) cm, 速生期分别为(34.1±0.39) cm、(0.269±0.006) cm。试验设4种处理:即对照、1次干湿交替(干旱—复水,A处理)、2次干湿交替(干旱—复水—干旱—复水,B处理)和3次干湿交替(干旱—复水—干旱—复水—干旱—复水,C处理),每处理3个重复(即3盆苗)。对照始终保持含水量在田间持水量的85%左右,其他在处理前先充分浇水3 d后任其自然干旱,以土壤含水量达到田间持水量的60%(中度胁迫)作为1次干旱标准,用称重法控制含水量(毛达如,1994),每次干旱后复水2 d进入下一次处理。同样的处理设2组(各12盆),一组为生理指标测定组,分别于各次处理干旱期末和复水后24 h取样测定幼苗生理指标;另一组为生长指标测定组,于处理前后测定幼苗高和地径生长,生长结束后进行生物量测定。
1.3 主要生理指标测定方法 1.3.1 光合速率、蒸腾速率在室外自然光照下测定,为减少每次测定时自然环境因子误差,测定时间均控制在9:00—10:00。每处理每重复选中等成熟的3片叶测定,仪器为美国产CI-301PS光合测定仪。
1.3.2 叶片质膜相对透性、叶片水分饱和亏缺、木质部水势叶片质膜相对透性采用电导率法,用DDB-6200型数字电导仪测定;叶片水分饱和亏缺(WSD)测定参照谢寅峰等(1999)的方法;用ZLZ-5型压力室测定幼苗主枝水势(长度10 ~15 cm)和主根水势。考虑到水势变化快,复水前后都在清晨7:00—8:00测定。
1.4 主要指标计算方法 1.4.1 水分胁迫指数(Wsi)水分胁迫指数反映胁迫条件下各指标偏离对照的程度,可用来反映幼苗受水分胁迫的影响程度,计算公式:Wsi=(XCK - Xi)/XCK,式中:Wsi为水分胁迫下某指标的水分胁迫指数;Xi为水分胁迫下某指标的测定值;XCK为对照相应指标的测定值。文中水分饱和亏缺、叶片质膜透性、根与茎木质部水势等指标与苗木抗旱性成负相关,取其倒数计算。Wsi值越大,表明该指标受水分胁迫的影响越大。
1.4.2 恢复度(Rd)恢复度是指水分胁迫解除后,各指标恢复情况与对照的贴近度。用下列公式计算:Rd=1-(XCK-Xr)/XCK,式中:Rd为水分胁迫解除后某指标的恢复度;Xr为水分胁迫解除后某指标测定值;XCK为对照相应指标的测定值。文中水分饱和亏缺、叶片质膜透性、根和茎木质部水势等指标与苗木抗旱性成负相关,取其倒数计算。Rd变化在0~1之间,其值越大,表明胁迫解除后,该指标与对照的贴近度越高,该指标恢复情况越好。
2 结果与分析 2.1 土壤干湿交替对不同生长时期青檀幼苗生理指标的影响 2.1.1 水分饱和亏缺从表 1看出,土壤干湿交替对青檀幼苗叶片水分饱和亏缺的影响程度因干湿交替处理次数、处理时期不同而异。生长初期幼苗的水分饱和亏缺(WSD)以A处理最大(比对照增加3.6倍),B处理WSD明显降低,C处理又呈上升趋势。这反映了生长初期幼苗对干湿交替环境从反应—适应—不适应的过程。速生期幼苗WSD随处理次数增加而逐渐增大,但A、B处理幼苗WSD比较接近,C处理因伤害加重,WSD急剧增加。方差分析表明,不同时期不同处理间WSD差异显著。幼苗水分胁迫指数(Wsi)与各时期WSD的变化规律一致。除A处理外,B、C处理Wsi值均表现为速生期高于生长初期,这是因为速生期幼苗生长旺盛,蒸腾作用强,再加上夏季高温低湿和土壤干旱的交互作用,对幼苗的伤害大于生长初期。
|
|
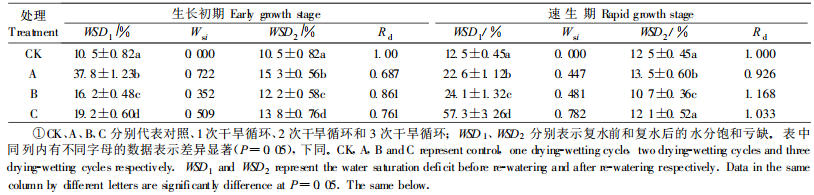
复水24 h后,幼苗叶片WSD得到不同程度的恢复, 但其恢复度(Rd)因干湿交替处理次数和处理时期而异。2个生长时期幼苗WSD的Rd值均表现为:B>C>A,这种变化与复水前WSD的变化基本吻合。A处理没经过适应性锻炼,复水24 h后恢复较慢, 而B处理幼苗经历了1次干旱锻炼,产生了一定的适应性,复水后恢复迅速。幼苗的适应性变化是有一定限度的,超过其限度,其恢复调节能力减弱。因此,C处理幼苗WSD的恢复能力小于B处理。表列数据还显示,复水后,速生期幼苗WSD的Rd大于生长初期,表明随幼苗年龄的增大对反复干旱的适应能力增强。此外,在速生期经过2次、3次干旱复水后的Rd值均超过了对照,这可以看作是复水后的一种补偿现象。
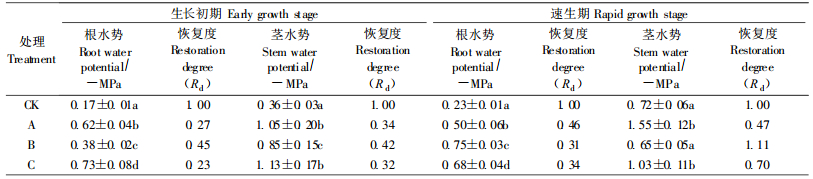
2.1.2 根、茎木质部水势从表 2可知:生长初期幼苗根木质部水势随处理次数增加而降低,茎木质部水势在3次处理中比较接近,但却明显低于对照; 速生期幼苗根、茎木质部水势均随处理次数的增加而逐渐降低。2个生长时期幼苗的根、茎木质部水势差均以A处理最大,B、C处理与对照接近。同一次处理,根木质部水势的Wsi总是大于茎木质部,速生期根茎水势的Wsi小于生长初期。说明干湿交替环境下,根木质部水势受到的影响大于茎木质部水势,生长初期大于速生期。
|
|
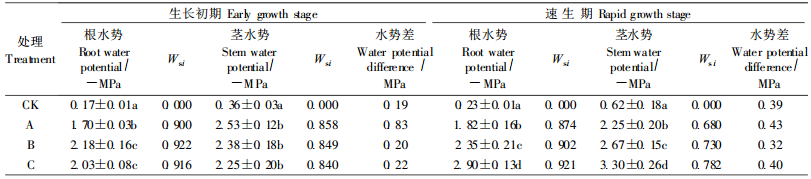
复水24 h后,幼苗根、茎木质部水势都得到不同程度的恢复(表 3)。生长初期幼苗根、茎水势,速生期幼苗茎水势均以B处理的Rd最大,速生期的Rd甚至超过1,而A、C处理Rd相对较小。速生期根水势的Rd值则是A处理最大,B、C处理比较接近。从表中还可看出,幼苗茎水势的Rd大于根水势,这有利于增大水势差,增强吸水能力,是幼苗适应干旱胁迫的策略。总体上,复水后速生期幼苗根、茎水势的Rd大于生长初期,表明速生期幼苗对反复干旱的适应能力强,受伤后自我恢复的能力强。
|
|
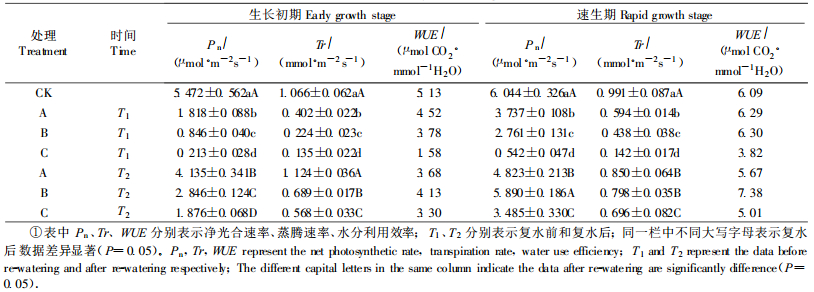
试验结果(表 4,表 5)表明,干湿交替处理下幼苗净光合速率(Pn)、蒸腾速率(Tr)变化的总趋势是随着干湿交替处理次数的增加而逐渐降低,其对应的Wsi值逐渐增大。就不同处理时期而言,生长初期Pn、Tr指标的Wsi均大于速生期,也反映出速生期幼苗忍耐反复干旱的能力强于生长初期。复水后,生长初期幼苗的Pn、Tr值和其对应的Rd值均随处理次数增加而逐渐减小,而速生期幼苗的Pn、Tr值和其Rd值的变化则没有明显的规律性。
|
|
|
|
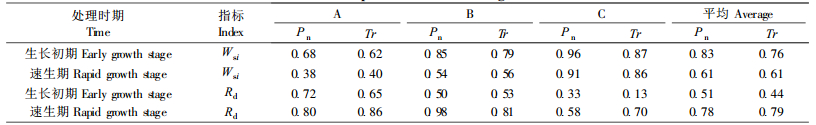
从表 4可看出,生长初期,幼苗的WUE随处理次数的增加而降低,速生期则表现为1次处理和2次处理WUE明显高于对照,而3次处理则低于对照。说明速生期幼苗在1~2次干湿交替环境下有助于提高WUE,而多次的干湿交替处理则会降低WUE。复水后,除1次处理外,其他处理的WUE都明显高于复水前,说明多次处理复水后植物的水分利用效率提高。
2.1.5 叶片质膜相对透性不同干湿交替处理幼苗叶片的质膜透性均明显大于对照(图 1), 而不同处理间叶片质膜透性的变化差异不大,尤其在速生期变化较小。表明每次处理之间的复水过程大大减轻了干旱胁迫对幼苗质膜的进一步伤害。生长初期1~3次处理叶片质膜透性的Wsi值分别为0.450、0.413和0.512,速生期分别为0.257、0.301和0.334,说明干湿交替处理对生长初期幼苗质膜透性的影响大于速生期。

|
图 1 不同干湿交替处理下幼苗叶片质膜透性变化 Fig. 1 The variation of plasm membrane permeability of seedlings under different drying-wetting cycles condition |
试验结果表明,干湿交替处理期间,2个生长时期幼苗高净生长量均随处理次数增加而逐渐降低(表 6),速生期高生长受水分胁迫的影响大于生长初期。地径净生长量在生长初期各处理之间变化没有明显规律性,且处理间差异不大,而速生期则表现为随处理次数增加而逐渐降低,但速生期地径净生长量小于生长初期。从Wsi值变化可看出,反复干旱胁迫对幼苗高生长量的影响大于地径生长量,其原因是水分胁迫对细胞伸长的抑制大于对细胞分裂的抑制(克雷默尔,1989),水分胁迫抑制顶端分生组织的延伸及分化,因而高生长受到抑制。由此可以推断,反复干旱胁迫对植物高生长的强烈抑制是喀斯特森林树种生长较慢而平均高度相对较低的原因之一。不同处理对幼苗主根长度的影响没有明显规律性。
|
|
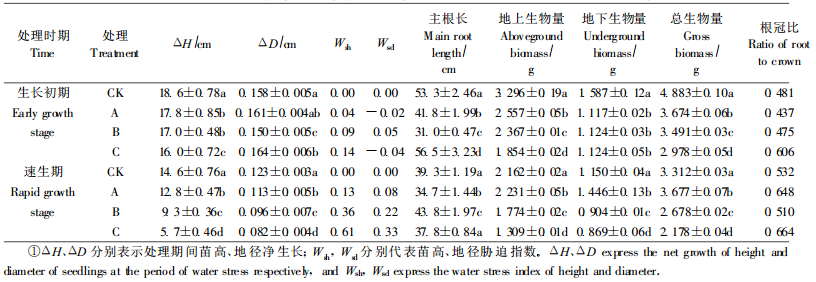
幼苗地上、地下部分生物量和总生物量变化总的趋势是随着干湿交替处理次数的增加而降低,速生期幼苗各生物量指标都小于生长初期。因为生长初期胁迫解除后,幼苗即进入速生期,此时外界环境、幼苗的生理状况都有利于生长;而速生期胁迫解除后,幼苗错过了生长的最佳时期,因而生长量较低。此外,干湿交替处理还影响幼苗的生物量分配, 随着处理次数的增加,根冠比有增加趋势,表明干湿交替处理有助于增加地下部分生物量。
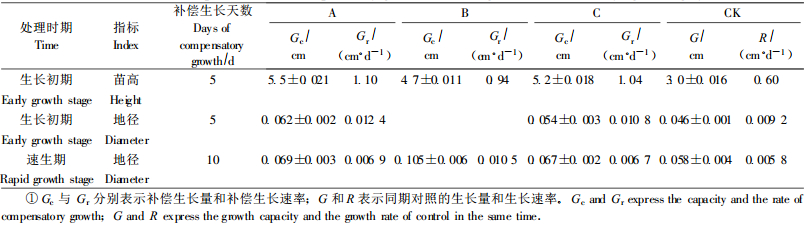
2.2.3 干旱复水后的补偿生长试验结果表明,干湿交替处理复水后,青檀幼苗高、地径生长普遍存在明显超过对照的补偿生长现象(表 7),但补偿生长的时间、补偿生长量和补偿生长速率因处理次数、处理时期不同而异。苗高补偿生长只在生长初期处理后出现,而地径在2个生长期均出现补偿生长,且速生期幼苗的地径补偿生长量大于生长初期,这与苗木的高、地径生长节律有关。青檀幼苗在旱后复水的补偿生长在一定程度上弥补了胁迫造成的损失,是该树种长期适应干湿交替环境的策略。
|
|
研究表明,干湿交替处理下青檀幼苗各项生理及生长指标的变化本质上与前人(韦小丽等,2005;侯常英等,2003)的研究一致,表现为随干湿交替处理次数的增加,水分饱和亏缺和质膜透性增加,水势、净光合速率、蒸腾速率和各项生长指标降低。但因每次处理后的短暂复水,使幼苗得到不同程度的恢复,一些指标如水势、WSD的变化相对缓和。复水后青檀幼苗各指标的恢复度与干湿交替处理次数呈负相关,幼苗经历的干旱次数越多,复水恢复的难度越大。干湿交替环境下,青檀幼苗的适应途径多样,如水分胁迫下通过保持相对稳定的水势差促进根系吸水,保持体内水分平衡;将更多的资源分配到地下部分,扩大根系以增大对水肥的吸收面积和复水后补偿生长等。
3.2 关于复水后的补偿效应许多研究表明作物对干旱-复水这种变水条件的响应方式是在胁迫解除后存在补偿效应,并认为这是作物对环境变化的一种适应,适应的结果体现在高度、叶面积、生物量、恢复生长的速率等方面的变化上(刘永红等,2006;黄占斌,2000;胡田田等,2005)。干旱复水后青檀幼苗的补偿效应普遍存在,表现在生理和生长两方面。如增大水势差,降低蒸腾速率、提高水分利用效率,最终苗高、地径均出现了补偿生长现象,但补偿生长持续的时间、补偿生长量、补偿生长速率与幼苗的生育期和不同指标的生长节律有关,在前人的研究中木本植物缺少这方面的报道。在干旱-复水后青檀幼苗表现出的这种补偿效应在生产上有极其重要的意义,生长初期幼苗可通过反复的干湿交替处理进行抗旱锻炼,既有利于增强抗旱性,又因补偿效应的存在而对生长的影响不大,速生期幼苗则应尽量避免反复的干旱-复水。
3.3 关于植物抗旱性研究方法的考虑植物生存的自然环境是一种多变水环境,植物与水分的关系在长期协调中达成一种内外系统的动态平衡,形成了相应的反应和适应机制。但传统的植物抗旱性研究习惯于用静态的和标准的干湿对比法,以至得出的结论有一定的局限性。农业科技工作者率先打破了这一传统,对反复干旱-复水条件下作物的生理生化基础进行了诸多研究(山仑等,2000;梁宗锁等,2001;Marja et al., 2005; Tezara et al., 2002),且有了新的发现,即干旱-复水的补偿效应,为节水农业提供了新的生物学思考。关于木本植物对干湿交替环境的反应和适应的研究报道较少,国内仅李吉跃(1999)对多重复干旱循环下苗木的气体交换和水分利用效率进行过研究,今后应加强这方面的研究。
程孝霞, 崔同林. 2005. 青檀壮苗培育. 林业实用技术, (2): 21-22. |
丁佐龙, 何云核, 高慧, 等. 1997. 青檀叶营养成分分析. 安徽农业大学学报, 24(10): 18-20. |
方升佐, 李光友, 李同顺, 等. 2001. 经营措施对青檀人工林生物量及檀皮质量的影响. 南京林业大学学报, 25(1): 21-25. |
方升佐, 李光友, 洑香香, 等. 2002. 立地条件对青檀檀皮中矿质元素含量的影响. 林业科学, 38(1): 8-14. DOI:10.3321/j.issn:1001-7488.2002.01.002 |
洑香香, 方升佐, 杜艳. 2002. 青檀种子休眠机理及发芽条件探讨. 植物资源与环境学报, 11(1): 9-13. DOI:10.3969/j.issn.1674-7895.2002.01.003 |
侯常英, 方升佐, 薛建辉, 等. 2003. 干旱胁迫对青檀等树种苗木生长及生理特性的影响. 南京林业大学学报, 27(6): 103-106. DOI:10.3969/j.issn.1000-2006.2003.06.025 |
胡田田, 康绍忠. 2005. 植物抗旱性的补偿效应及其在农业节水中的应用. 生态学报, 25(4): 885-891. DOI:10.3321/j.issn:1000-0933.2005.04.034 |
黄占斌. 2000. 干湿变化与作物补偿效应规律研究. 生态农业研究, (3): 30-33. |
克雷默尔.1989.植物的水分关系.许旭旦, 汤章城, 王万里, 等译.北京: 科学出版社, 470-471
|
李吉跃. 1999. 多重复干旱循环对苗木气体交换和水分利用效率的影响. 北京林业大学学报, 21(3): 1-8. DOI:10.3321/j.issn:1000-1522.1999.03.001 |
刘永红, 杨勤, 杨文钰, 等. 2006. 花期干湿交替对玉米干物质积累与再分配的影响. 作物学报, 32(11): 1723-1727. DOI:10.3321/j.issn:0496-3490.2006.11.021 |
梁宗锁, 康绍忠, 韶明安, 等. 2001. 土壤干湿交替对玉米耗水特性及水分利用的影响. 土壤学报, 38(3): 391-39. |
毛达如. 1994. 植物营养研究法. 北京: 中国农业大学出版社, 10-11.
|
山仑, 苏佩, 郭礼昆, 等. 2000. 不同类型作物对干湿交替环境的反应. 西北植物学报, 20(2): 164-170. DOI:10.3321/j.issn:1000-4025.2000.02.002 |
韦小丽, 徐锡增, 朱守谦. 2005. 水分胁迫下3个榆科树种幼苗生理生化指标的变化. 南京林业大学学报, 29(2): 47-50. |
谢寅峰, 沈惠娟, 罗爱珍, 等. 1999. 南方7个造林树种幼苗抗旱生理指标比较研究. 南京林业大学学报, 23(4): 13-16. DOI:10.3969/j.issn.1000-2006.1999.04.003 |
朱守谦. 2003. 喀斯特森林生态研究(Ⅲ). 贵阳:贵州科技出版社: 12-13. |
Fang Shengzuo, Song Liyi, Fu Xiangxiang. 2006. Effects of NaCl stress on seed germination, leaf gas exchange and seedling growth of Pteroceltis tatarinowii. Journal of Forestry Research, 17(3): 185-188. DOI:10.1007/s11676-006-0043-z |
Marja A van, Staalduinen, Niels P R, et al. 2005. Differences in the compensatory growth of two co-occurring grass species in relation to water availability. Oecologia, 146: 190-199. DOI:10.1007/s00442-005-0225-y |
Tezara W, Mitchell V, Driscoll S P, et al. 2002. Effects of water deficit and its interaction with CO2 supply on the biochemistry and physiology of photosynthesis in sunflower. Journal of experimental Botany, 53(375): 1781-1791. DOI:10.1093/jxb/erf021 |