 2007, Vol. 43
2007, Vol. 43文章信息
- 郭蓓, 刘勇, 李国雷, 甘敬, 徐扬.
- Guo Bei, Liu Yong, Li Guolei, Gan Jing, Xu Yang.
- 飞播油松林地土壤酶活性对间伐强度的响应
- Response of Soil Enzyme Activity to Thinning Intensity of Aerial Seeding Pinus tabulaeformis Stands
- 林业科学, 2007, 43(7): 128-133.
- Scientia Silvae Sinicae, 2007, 43(7): 128-133.
-
文章历史
- 收稿日期:2006-12-21
-
作者相关文章



2. 北京市园林绿化局 北京 100029
2. Beijing Municipal Bureau of Landscape and Forestry Beijing 100029
森林土壤酶系统是森林土壤中生物活动的产物,其活性受森林土壤理化性质、植被种类和生物多样性等生态因子的综合影响(关松荫,1986;杨承栋等,1999;杨万勤等,2004)。目前普遍认为森林土壤酶系统是森林生态系统的物质循环和能量流动等生态过程中最为活跃的生物活性物质,在森林生态系统的关键过程中扮演着重要的角色(周礼恺,1987)。因此,土壤酶活性可以被用作土壤生态胁迫或土壤生态恢复等早期的敏感性指标(Dick et al., 1993; Garcia et al., 1997)。最近研究表明,选择适宜间伐可改善林地水热状况,增加林下植物多样性,提高微生物活性(杨万勤,2004)。因此探讨间伐强度-林下生物多样性-土壤酶活性三者之间的关系,对深入理解生态系统过程具有重要意义,但迄今为止,尚无有关此方面的报道。
飞播造林是人工促进森林进展演替而形成的人工经营的自然体系( 《中国飞播造林四十年》编委会,1998),是偏远山区重要植被恢复模式之一。飞播油松林的形成是由飞籽侵入荒山灌丛群落、竞争、定居的过程,其群落类型较为特殊(陈芳清等,1997),种群空间格局为聚集度较高的群团型(刘建军等,1991;刘向东等,1983),密度普遍较大,郁闭相对迅速。针叶林由于密度过高引发的土壤退化问题已受到广大学者关注(孙书存等,2005),选择适宜间伐强度是飞播林健康生长和发挥最大生态效益的关键措施(郭天亮等, 2004)。但目前的为数不多的一些关于飞播林的研究也还只是分析了保留密度对其生长的影响(杨澄,1996;莫冀翔等,2003;刘占朝等,2003;郭天亮等,2004),间伐强度的筛选仍以追求木材产量为目标,忽视了处于生态脆弱区飞播林生态效益的发挥。为此,本文以地位级Ⅳ、9年生的飞播油松幼林为研究对象,通过间伐5年后林下植被调查和土壤酶的测定,揭示间伐强度对幼龄油松林植被发育的影响,进而探讨林下植被对土壤酶活性的作用,从土壤酶活性响应差异性角度评判间伐强度的优劣,从而为飞播油松林的可持续经营提供理论依据。
1 研究地概况研究地点位于北京市延庆县王家堡飞播造林基地,地理位置为40°16′ N,115°40′ E。地貌属低山丘陵,海拔600~1 200 m。气候属暖温带大陆性季风气候。年平均气温8.8 ℃,平均降水量450~550 mm。土壤类型为含石砾较多的山地褐土,成土母岩以花岗岩为主。
1990年初营造的飞播油松林,面积600 hm2,有效面积400 hm2,坡位中,坡向北坡。2000年进行了4种不同间伐强度的处理:Ⅰ为弱度间伐、Ⅱ为中度间伐、Ⅲ为强度间伐、Ⅳ为超强度间伐。保留密度分别为5 600, 4 100, 3 700,1 830株·hm-2,间伐后即进行封山育林,林下植被的生长发育很少受人为干扰。2005年3月初,在上述不同处理的油松林中,分别选定20 m×15 m的4块有代表性的试验地作为样地林分,同时设置初植密度为8 000株·hm-2的样地作为对照(CK)。样地资料见表 1。5样地林下植被优势种分别为土庄绣线菊(Spiraea pubescens)+多花胡枝子(Lespedeza floribunda)-京芒草(Achnatherum pekinense)+深山堇菜(Viola selkirkii)、荆条(Vitex negundo)+土庄绣线菊-紫花地丁(Viola yedoensis)+白莲蒿(Artemisia gmelinii)、土庄绣线菊+三裂绣线菊(Spiraea trilobata)-野青茅(Calamagrostis arundinacea)+山莴苣(Lactuca indica)、荆条+土庄绣线菊-披针叶苔草(Carex lanceolata)+鼠掌老鹳草(Geranium sibiricum)、虎榛子(Ostryopsis davidiana)+土庄绣线菊-野古草(Arundinella hirta)+京芒草。
|
|
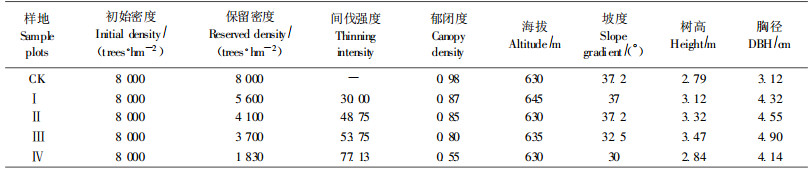
2005年8月初分别在5样地中以对角线方式设置5个样点,设立土壤剖面,按0~20 cm,20~40 cm,40~60 cm土层深度采样(分别记为A,B,C层),去除石块、根系和土壤动物,把每样地同一层次的土样等量混合均匀。取回的土壤样品经风干后,碾碎过筛,贮存于广口瓶中,4 ℃储藏备用。
2.2 林下植被调查2005年8月初,在每个样地内对乔木每木检尺,调查乔木胸径、高度等,同时记载样地的海拔、坡向、坡度等环境因子。每个样地内沿对角线设置5 m×5 m的样方5个,按常规分别记载灌木和草本的种类、高度、数量和盖度等。同时,在每块固定样地内,机械均匀设置1 m×1 m样方10个,调查林下各种草本植物的种类。
2.3 土样分析土壤理化性质的测定,采用《土壤农化分析》(鲍士旦, 2005)上的方法,土壤含水量的测定采用烘箱法,有效磷采用氟化铵-盐酸浸提法,速效钾采用乙酸铵浸提-火焰光度法,有机质采用重铬酸钾氧化-外加热法,pH值采用电位法。土壤酶活性的测定采用关松荫(1986)方法,过氧化氢酶采用高锰酸钾滴定法,多酚氧化酶采用碘量滴定法,碱性磷酸酶采用磷酸苯二钠比色法,脲酶采用靛酚比色法,转化酶采用硫代硫酸钠滴定法。
2.4 数据处理Shannon-Wiener指数(H)=-∑PilnPi;Pielou指数(J)=H/lnS。式中:S为物种数目,A为样方面积,Pi为种i的相对重要值(马克平等,1995;毕润成,1999)。
试验数据采用SPSS统计软件分析。双变量相关分析采用Pearson相关系数。
3 结果与分析 3.1 土壤酶活性的剖面分布特征由表 2可知,过氧化氢酶在各土层间变化不明显,脲酶、碱性磷酸酶、转化酶、多酚氧化酶等活性随土层的增加而减小,表现出较强的褐土酶特性(关松荫,1986)。与对照相比,间伐可提高土壤酶活性,B、C层酶活性所受影响较小,两层酶活性趋近,A层酶活性受干扰最大,随间伐强度的增大,A层5种土壤酶活性升高的程度表现不一,脲酶、碱性磷酸酶活性弱度间伐最大,中度间伐次之,而转化酶、多酚氧化酶活性却呈相反的变化趋势。间伐后土壤多酚氧化酶活性的提高将有助于促进土壤的酚类物质向醌类物质的转化,这对于减少土壤酚类物质的累积,降低土壤毒素的含量有着积极的意义(张鼎华等,2001)。
|
|
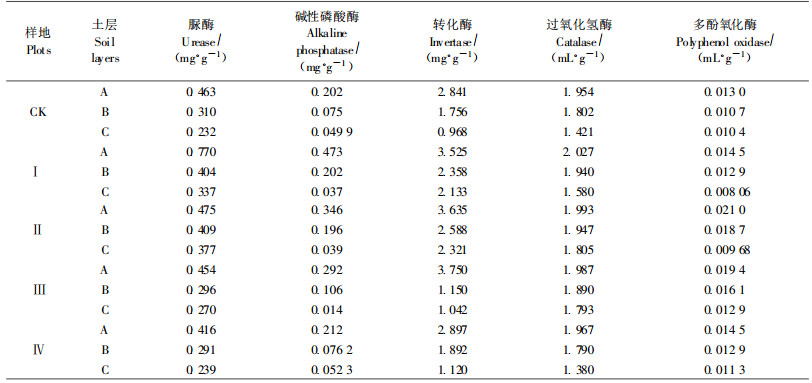
由表 3可知,土壤含水量、pH值、土壤养分随土壤层次的加深而较小。土壤剖面中,A层受间伐的影响最大,这与土壤酶活性表化趋势基本一致。与对照相比,A层土壤含水量、全氮、速效钾、有效磷、有机质都有不同程度的增加,而pH值对间伐的响应相对于土壤理化性质要复杂得多,弱度、中度、强度间伐土壤A层pH值均小于对照,仅有超强度间伐样地有一定增加。
|
|
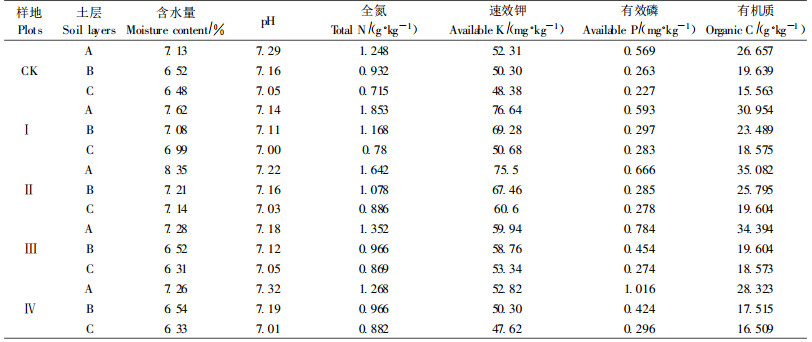
养分含量的变化与酶活性是密切相关在许多研究中已经被证实(孙庆业等,2005;Aon et al., 2001;Ros et al., 2003),但间伐过程中土壤酶活性与土壤养分的定量分析尚未见报道。由表 4可知,速效钾、有机磷、有机质、全氮均与土壤酶正相关,其中全氮和有机质与5种土壤酶关系最为密切,均达到极显著正相关。间伐使林地土壤有机质含量的增加,这既能增强土壤孔隙度和通气性,又能为微生物提供丰富的营养源,有利于微生物、土壤酶和矿物质的固定。可见,作为土壤酶的有机载体,有机质对土壤酶的影响较无机态的速效钾、有效磷明显。值得注意的是,碱性磷酸酶与速效钾极显著正相关,而与有效磷正相关,有驳于土壤磷酸酶活性对磷的依赖强于钾的观点(关松荫,1986),这可能与土壤类型或林地土壤K/P营养元素失调有关。
|
|
由表 5可知,与对照相比,弱度、中度、强度间伐可增加林下植被多样性,其中以中强度间伐效果最为明显。而超强度间伐Ⅳ林下植被的Shannon-Wiener、Pielou指数下降。
|
|
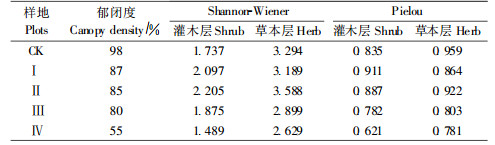
由表 6可知,土壤脲酶、碱性磷酸酶、转化酶、过氧化氢酶等酶活性与林分郁闭度、灌木层和草本层的Shannon-Wiener、Pielou指数成正相关,其中碱性磷酸酶与郁闭度、林下植被多样性关系最为密切。进而分析林分郁闭度与灌木层和草本层的Shannon-Wiener、Pielou指数,发现两者成正相关或极显著正相关,这表明间伐通过减小高密度飞播油松的郁闭度,不仅增加了林下灌木和草本植物的种类和多样性,而且使林下植被的优势种各异,林下植物可通过根系分泌物和凋落物质量等影响土壤酶活性(杨万勤等,2004)。
|
|
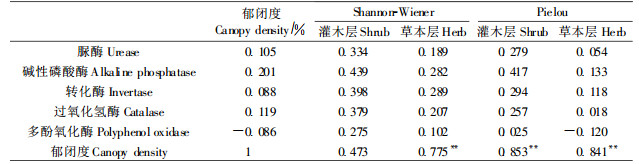
多酚氧化酶与林分郁闭度呈负相关,与灌木层和草本层Shannon-Wiener指数、灌木层Pielou指数正相关,仅与草本层Pielou指数负相关,因此,多酚氧化酶与郁闭度的负相关可能是与由草本层分布不均所致。
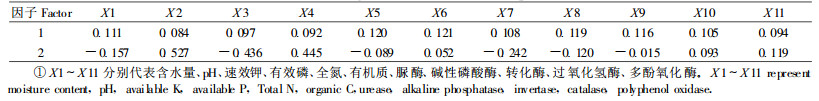
3.4 土壤质量的评价指标分析为了更好的分析不同土壤层次的酶活性与土壤理化性质的关系,从而选择用于飞播油松林土壤健康评价指标,对研究区域内的土壤信息系统的土壤酶、土壤含水量、土壤养分和pH值共11个指标进行了主成分分析(表 7)。需2个主成分即可解释超过83%的信息。由表 8可知,第一主成分为全氮、有机质、转化酶、过氧化氢酶;第二主成分为含水量、pH、速效钾、有效磷、脲酶、碱性磷酸酶、多酚氧化酶。说明土壤的养分中全氮和有机质的含量多少是评价土壤质量的重要指标,土壤酶中转化酶和过氧化氢酶的活性对土壤质量的高低评价有重要影响。
|
|

|
|
飞播林群落代替荒山宜林群落而加快了演替进程,同时也加大了对土壤养分的消耗。飞播林密度普遍较大,养分竞争激烈,加之飞播林以针叶纯林为主,郁闭度大,林下光照差,土壤微生物较少(周德明等,2002;李志辉等,2004),凋落物分解缓慢,养分返还少,飞播林“取”多“给”寡,林地肥力下降,因此,油松飞播林郁闭后应及时进行间伐。在土壤理化性质、土壤酶及pH值构成的土壤质量评价指标体系中,只需2个主成分即可解释超过83%的信息。5种土壤酶对间伐敏感性各异,碱性磷酸酶受间伐影响最大,脲酶次之,转化酶、多酚氧化酶居中,过氧化氢酶所受影响最小;不同层次的土壤酶对间伐响应也不同,A层土壤酶活性变化最为明显,随土层深度的增加,酶活性趋同且对间伐敏感性减弱。造成这种原因,笔者认为主要有以下几方面因素:一方面,土壤养分随土壤层次的加深而减小,而土壤酶主要是以物理的或化学的结合形势吸附在土壤有机质或无机颗粒上,或与腐殖质络合。土壤有机质是土壤中各种营养元素特别是氮、磷的主要来源,它能使土壤具有保肥力和缓冲性能使土壤疏松,从而改善土壤理化性质。本试验中的5种酶活性与速效钾、速效磷存在着不同程度的正相关,均与有机质、全氮含量极显著正相关,也说明了作为土壤酶有机载体的有机质固定土壤酶的作用强于速效钾、速效磷等无机物质。另一方面,间伐有效调节了林分的郁闭度,飞播油松植被优势种各异,改变了根系的组成,根系种类和数量增大;同时,间伐使灌木层、草本层的多样性和均匀性也有一定程度的提高,由于灌木和草本层根系分布较浅,并且多为细根,间伐影响了根系的分布。细根是根系最活跃的部分,其分泌物和细胞的脱落促使酶进入土壤,而细根也主要分布在土壤的表层,在土壤中由上而下逐渐减少。再加上随着土层的增加,通气状况越来越差,微生物种类和数量递减,因此,土壤酶活性随土壤深度的增加逐渐减弱(关松荫等,1984),我们通过土壤酶活性与林分郁闭度、林下植被的多样性和均匀性的相关分析有力证明了这一推断。
及时对郁闭林分进行适宜强度的间伐是提高飞播油松林地土壤酶活性的有效途径。中度间伐下5种土壤酶活性普遍较高,结合土壤的理化性质及其林下植被物种组成、多样性指标,地位级Ⅳ、初始密度为8 000株·hm-2、9年生飞播油松幼林选择48.75%的间伐强度最佳。飞播油松林多处于海拔较高、交通不便的深远山区,林分高密度过大,可适当增大飞播林间伐强度从而延长间伐周期以达到减少间伐次数的目的,本研究认为可把上述条件的林分密度降至3 700株·hm-2左右。而保留密度1 830株·hm-2的林分,林下植被组成单一,土壤酶活性与对照相差不大,超强间伐强度在生产实践中要慎重使用。
《中国飞播造林四十年》编委会. 1998. 中国飞播造林四十年. 北京: 中国林业出版社.
|
鲍士旦. 2005. 土壤农化分析. 北京: 中国农业出版社.
|
毕润成. 1999. 山西霍山山核桃群落生态特征及其区系分析. 应用生态学报, 10(6): 650-656. |
陈芳清, 卢斌. 1997. 长江飞播油松林的群落学特点. 信阳师范学院学报, 10(3): 42-46. |
关松荫, 沈桂琴, 孟昭鹏, 等. 1984. 我国主要土壤剖面酶活性状况. 土壤学报, 21(4): 368-381. |
关松荫. 1986. 土壤酶及其研究法. 北京: 农业出版社, 188-359.
|
郭天亮, 李素林, 白静, 等. 2004. 飞播油松林抚育间伐技术的探讨. 防护林科技, (5): 63-64. |
李志辉, 漆良华, 柏方敏, 等. 2004. 马尾松飞播林土壤肥力研究. 中南林学院学报, 24(5): 32-35. DOI:10.3969/j.issn.1673-923X.2004.05.007 |
刘建军, 杨澄, 吴成儒. 1991. 油松飞播林空间格局分析. 西北林学院学报, 6(2): 51-54. |
刘向东, 吴钦孝, 侯庆春, 等. 1983. 飞播油松幼苗的生态分布. 水土保持通报, (6): 77-82. |
刘占朝, 王团荣, 张宏文, 等. 2003. 河南省油松飞播林林分密度控制图编制. 林业资源管理, (4): 24-27. DOI:10.3969/j.issn.1002-6622.2003.04.007 |
马克平, 黄建辉, 于顺利. 1995. 北京东灵山地区植物多样性的研究. 生态学报, 15(3): 268-277. DOI:10.3321/j.issn:1000-0933.1995.03.006 |
莫翼翔, 孙丙寅, 康克功, 等. 2003. 保留密度对油松飞播林生长的影响. 西北林学院学报, 18(4): 57-59. DOI:10.3969/j.issn.1001-7461.2003.04.018 |
孙庆业, 任冠举, 杨林章, 等. 2005. 自然植物群落对铜尾矿废弃地土壤酶活性的影响. 土壤学报, 42(1): 37-43. DOI:10.3321/j.issn:0564-3929.2005.01.006 |
孙书存, 包维楷. 2005. 恢复生态学. 北京: 化学工业出版社, 50.
|
杨澄. 1996. 油松飞播林生长规律的研究. 西北林学院学报, 11(4): 24-27. |
杨承栋, 焦如珍, 屠星南, 等. 1995. 杉木林下植被对5~15 cm土壤性质的改良. 林业科学研究, 8(5): 514-519. |
杨万勤, 王开运. 2004. 森林土壤酶的研究进展. 林业科学, 40(2): 152-159. DOI:10.3321/j.issn:1001-7488.2004.02.027 |
张鼎华, 叶章发, 范必有, 等. 2001. 抚育间伐对人工林土壤肥力的影响. 应用生态学报, 12(5): 672-676. DOI:10.3321/j.issn:1001-9332.2001.05.007 |
周德明, 陈晓萍, 张建湘, 等. 2002. 马尾松飞播林地土壤微生物的研究. 中南林学院学报, 22(3): 59-62. DOI:10.3969/j.issn.1673-923X.2002.03.009 |
周礼恺. 1987. 土壤酶学. 北京: 科学出版社, 178.
|
Aon M A, Colaneri A C. 2001. Temporal and spatial evolution of enzyme activities and physio-chemical properties in an agricultural soil. Applied Soil Ecology, 18: 255. DOI:10.1016/S0929-1393(01)00161-5 |
Cookson P, Lepiece A G. 1996. Urease enzyme activities in soils of the Batinah region of the Oman. Journal of Arid Enviroments, 32: 225-238. DOI:10.1006/jare.1996.0019 |
Dick W A, Tabatabai M A. 1993. Significance and potential uses of soil enzymes//Blaine Metting F. Soil Microbial Ecology. Application in Agricultural and Environmental Management. New York: Marcel Dekker, 95-127
|
Garcia C, Hernandez T. 1997. Biological and biochemical indicators in derelict soils subject to erosion. Soil Biol Biochem, 29(2): 171-177. DOI:10.1016/S0038-0717(96)00294-5 |
Ros M, Hermandez M T, Carcia C. 2003. Soil microbial activity after restoration of a semiarid soil by organic amendents. Soil Biology & Biochemistry, 35: 463-469. |