 2007, Vol. 43
2007, Vol. 43文章信息
- 王军, 韦爱梅, 孙思.
- Wang Jun, Wei Aimei, Sun Si.
- 青枯菌对桉树及非寄主树木根部吸附和侵入的比较
- Comparison on Adsorption and Ingression of Ralstonia solanacearum to Eucalyptus and a Nonhost Tree
- 林业科学, 2007, 43(7): 51-54.
- Scientia Silvae Sinicae, 2007, 43(7): 51-54.
-
文章历史
- 收稿日期:2005-12-16
-
作者相关文章


植物细菌病害的发生,始于病原细菌对寄主的吸附和接触,吸附的选择性或专化性反映在病原细菌与寄主之间的识别上(Lippincott et al., 1984)。识别可促进或抑制病原物的进一步活动,对于病原物是否能与寄主最终建立寄生关系起着重要作用(董汉松,1993)。在青枯菌(Ralstonia solanacearum)-植物病理系统中,一些研究就病原细菌对番茄(Lycopersicon esclentum)(王卉等,1993)和木麻黄(Casuarina equisetifolia)(罗焕亮等,2002)的吸附识别进行了初步试验,但在桉树(Eucalyptus)青枯病上,现有的研究主要集中在病害的发生规律和防治方面(王军等,1997;向妙莲等,2004;韦爱梅等,2004;孙思等,2004),对于病原细菌的吸附识别及侵染机制还少有涉及(王军,2005)。为了更深入地探讨桉树青枯病的发生机理,本研究通过比较研究青枯菌对桉树及非寄主树木根部的吸附及侵入的选择性,以了解该病理系统中病菌与寄主之间最初的互作情况,并揭示它们之间可能存在的识别现象。
1 材料与方法 1.1 供试材料供试细菌:青枯菌,大肠杆菌(Escherichia coli)JM109菌株,由华南农业大学林学院保存;分别培养青枯菌(TTC培养基)和大肠杆菌(NA固体培养基)24~48 h,挑取并用无菌水配制成浓度为3×108 cfu·mL-1的菌悬液(McFarland云度计)(方中达,1998)备用。
供试树木:青枯菌寄主尾巨桉(Eucalyptus urophylla × E. grandis) 196(感病),尾巨桉3229(抗病),由广东省林科院提供。非寄主红花羊蹄甲(Bauhinia blakeana)实生苗,由华南农业大学提供。各树苗苗高10~15 cm,生长健壮,其抗(感)病性均经实验室人工接种验证。
1.2 试验方法 1.2.1 苗木接种参照施仲美等(1998)的WFT方法,并对根部作微伤或无伤处理,进行青枯菌悬液水培接种;对照用无菌水代替青枯菌悬液,然后均置于人工气候箱培养(30 ℃,RH=70%~80%)。
1.2.2 细菌的计数采用血球计数法直接计数:取待测菌悬液于血球计数板上,在暗视野显微镜高倍镜下计数细菌个数,然后计算每毫升悬浮液细菌的数目(方中达,1998),并换算成“菌量(个)/根质量(g)”,即[细菌总数(mL-1)×分离用水量(mL)]/根质量。
1.2.3 根部菌量检测根表吸附菌量:分别于接种后2 h和24 h,在植株距根尖切口截取主根约2 cm左右,浸入10 mL无菌水中3 min,并不断晃动洗涤下植物根表所吸附细菌体, 以血球计数法检测细菌个数,用同样方法检查对照根表是否带菌;每1个时间为1个处理,同一处理设5个重复。
根内含菌量:依据王卉等(1993)的方法略加修改。分别于接种后2 h和24 h,在植株距根尖切口截取主根约2 cm左右,无菌水冲洗3次,灭菌滤纸吸干,称质量,于10 mL无菌水内用镊子夹碎,静置25 min。倍比稀释,以血球计数法检测细菌数量,用同样方法检查对照根内是否带菌;每1个时间为1个处理,同一处理设5个重复。
2 结果与分析 2.1 寄主桉树与非寄主树木根表吸附菌量的变化比较抗、感病桉树寄主与非寄主根表吸附菌量,接种后2 h,无论是有伤或无伤,非寄主红花羊蹄甲与抗性寄主3229根表吸附的菌量均较感病寄主196根表为少。接种后24 h,非寄主和抗性寄主的根表吸附菌量进一步下降,而感病寄主根表吸附菌量进一步上升,它们之间差距进一步扩大(表 1)。对照根表无细菌吸附。
|
|
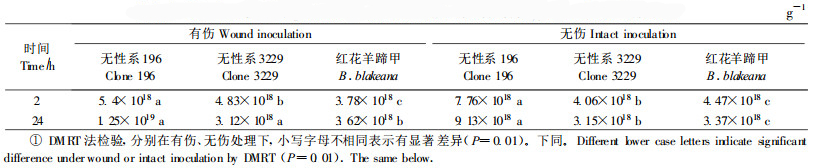
比较抗、感病桉树寄主与非寄主根内含菌量变化,结果表明:接种后2 h根内含菌量(细菌侵入量),无论是有伤或无伤,非寄主红花羊蹄甲明显较寄主为多,而抗病寄主3229根内含菌量却少于感病寄主196,不过接种后24 h,非寄主和抗病寄主3229根内含菌量均下降,而感病寄主196根内含菌量则显著上升,并都超过了非寄主与抗病寄主的根内含菌量(表 2)。对照根内未检查到细菌。
|
|

将青枯菌与非病原的大肠杆菌作对比测定,接种后2 h,青枯菌在感病寄主196根表的吸附量较大肠杆菌为多,有伤与无伤处理皆如此。接种后24 h,2种细菌的吸附量都有一定幅度的上升,但病菌的数量仍然多于非病原菌(表 3)。对照根表无细菌吸附。
|
|
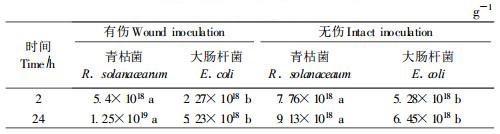
接种感病品系196后2 h,2种细菌在桉树根内的含量(细菌侵入量)有一定差异,无伤接种非病原菌的数量较多,有伤接种则病原菌较多。但接种后24 h,二者的数量均增加,有伤与无伤处理皆如此,不过,青枯菌的菌量增长更快(表 4)。对照根内未检查到细菌。
|
|

青枯菌对桉树的侵染,需要首先吸附到寄主的根表,然后进入根内组织。Marshall(1975)指出在接种细菌后的最初数分钟内,细菌便开始向寄主细胞表面聚集,借助纤维状物质与植物细胞壁相连,此时,用水或缓冲液极易洗脱,该阶段称为可逆性吸附阶段。一般情况下接种2 h以后,细菌多糖分子与植物细胞壁表面发生相应的化学反应,由生化力引起紧密的不容易用水洗脱的不可逆吸附。不可逆吸附通常由细菌的脂多糖(LPS)来完成的,而胞外多糖(EPS)则可使病原细菌不被吸附而游离并在寄主体内增殖(Graham et al., 1977)。
细菌对寄主植物根表的吸附量测定,以往偏重对不可逆吸附测定,通过撕去寄主根尖表皮而测量其含菌量(王卉等,1993;罗焕亮等,2002)。这种方法的前提是假定存在于表皮细胞的细菌全是处于不可逆吸附状态。但实际上由于EPS的作用,总有一部分细菌是不会被吸附而处于游离状态的。因此,这种对不可逆吸附的测定方法存在一定误差,而且要完整地撕去根尖部分的表皮也是一项非常困难的操作。
理论上虽然可以将细菌进入寄主植物细胞后的状态分为吸附与游离或增殖,但实际上要精确确定它们在根组织中的含量比例在目前的技术水平下是不大可能的,即要精确测定不可逆吸附的细菌吸附量是很难达到的。然而测定细菌对树木根表的可逆性吸附量,以及细菌对寄主根部的侵入量(本试验定义为接种后2 h时的根内含菌量)却是可行的。由于细菌在侵入根部后的初始阶段主要存在于表皮,细菌的侵入量与上述“表皮含菌量”有一定的一致性。吸附量(可逆性)和侵入量这2个指标,前者可以反映侵入前细菌对植物根表的相对选择性地聚集,后者则可以体现细菌对侵入树木根部的选择性。因此,通过测定二者在抗、感病寄主及非寄主上的差异来判断细菌与树木根部之间是否存在选择性或识别,比单一测定“表皮含菌量”所代表的不准确的不可逆吸附量具有更高的准确性。
细菌计数的常规方法一般采用平板涂布和显微镜下直接计数。前者得到的是菌落单位,由于单个菌落并不一定由单个细菌形成,且只能计测活的细菌,因而本试验选择了后者,利用暗视野显微镜在血球计数板上直接计数细菌的个数,这样可以更精确地得到细菌总的数量。
根据本试验结果,在吸附方面,青枯菌对感病寄主根表的吸附量较对抗病寄主和非寄主的吸附量多,且随时间差距扩大,表明感病寄主对病菌的亲和性较强;比较青枯菌与大肠杆菌,也是病原菌对寄主根表的吸附量大于非病原菌。因此,病原菌对寄主根表的吸附具有一定的选择性,且是亲和性的,这一点与早先在不可逆吸附上的研究结果认为是“逃逸性”的(Graham et al., 1977;董汉松,1991)不同。不过病原菌的这种选择性是微弱的,因为从绝对数量上看,无论是青枯菌对非寄主树木根表,还是非病原菌对桉树根表都存在相当可观的吸附量。
在侵入方面,青枯菌对感病寄主根部24 h的侵入量也多于对非寄主和抗病寄主根部的侵入量,表明病菌对树木根部的侵入也存在一定的选择性,且这种选择性是亲和性的。但与吸附一样,青枯菌对寄主根部侵入的选择性也是微弱的,因为病菌对抗、感病寄主和非寄主的侵入绝对量都相当大;比较青枯菌与大肠杆菌对感病桉树根部的侵入,甚至这种微弱的选择性也几乎不存在,这一结果与Saile等(1997)在番茄根部皮层曾观察到过的现象类似。
从统计学的角度看,青枯菌对寄主根部的吸附和侵入都显示了一定的选择性,并且在性质上是亲和性的。但是由于病菌能够大量地吸附和进入抗性寄主和非寄主根部,非病原细菌也能够大量地吸附和进入桉树根部,因此这种选择性的生物学意义不大。本试验结果至少在群体上不支持青枯菌对桉树根部有明显的接触性识别行为。病原细菌的吸附和侵入固然是病害得以发生的前提,但从本试验结果看青枯菌对桉树的侵染似乎并不依赖病菌对寄主初始的接触识别即吸附和侵入。由于无论是吸附量还是侵入量,在感病寄主上菌量都呈上升趋势,而在抗病寄主和非寄主上都呈下降趋势,因此,病菌吸附并侵入树木根内后的菌量变化趋势更有可能涉及或决定病害的最终发生与否,这点同早期的一些研究认为接触识别在启动或关闭侵染程序方面有重要作用的观点有所不同(董汉松,1993)。
董汉松. 1991. 植物识别子与病原细菌识别子间的相互作用. 植物生理学通讯, 27(5): 391-396. |
董汉松. 1993.寄主植物与病原微生物的相互识别//贾示荣.农业生物技术进展与展望——国家863计划生物技领域第一主题学术讨论会论文选编.合肥: 中国科学技术出版社, 59-65
|
方中达. 1998. 植病研究方法. 3版. 北京: 农业出版社, 182, 183, 190-191.
|
罗焕亮, 王军, 张景宁. 2002. 木麻黄青枯菌的致病性与其对根表吸附及在根内增殖关系的研究. 林业科学研究, 15(1): 21-27. DOI:10.3321/j.issn:1001-1498.2002.01.004 |
施仲美, 奚福生, 韦颖文, 等. 1998. 测定桉树品(种)系对青枯病菌抗性的技术研究. 广西林业科学, 27(4): 165-169. |
孙思, 韦爱梅, 伍慧雄, 等. 2004. 青枯病的化学与生物防治研究进展. 江西植保, 27(4): 157-162. DOI:10.3969/j.issn.2095-3704.2004.04.006 |
王卉, 任欣正. 1993. 青枯菌(Pseudomonas solanacearum)在番茄抗、感病品种根部的吸附、侵入和繁殖. 植物病理学报, 23(2): 143-150. DOI:10.3321/j.issn:0412-0914.1993.02.013 |
王军, 苏海, 岑炳沾. 1997. 林木青枯病研究综述. 华南农业大学学报, 18(4): 118-121. |
王军. 2005. 青枯菌对植物的致病机制及其调节. 林业科学, 41(3): 142-147. |
韦爱梅, 王军. 2004. 我国植物青枯病抗病性研究新进展. 广东林业科技, 20(4): 46-49. DOI:10.3969/j.issn.1006-4427.2004.04.013 |
向妙莲, 冉隆贤. 2004. 桉树青枯病研究进展. 中国森林病虫, 23(1): 37-40. DOI:10.3969/j.issn.1671-0886.2004.01.014 |
Graham T L, Sequeira L, Huang T R, et al. 1977. Bacterial lipopolysaccharides as inducers of disease resistance in tobacco. Applied and Environmental Microbiology, 34(4): 424-432. |
Lippincott J L, Lippincott B B. 1984. Concepts and experimental approaches in host-microbe recognition//Kosuge T, Nester E W. Plant-Microbe Interactions, Molecular and Genetic Perspectives: Vol. 1. MPC, New York, CMP, London, 195
|
Marshall K C. 1975. Clay mineralogy in relation to survival of soil bacteria. Annual Review of Phytopathology, (13): 357-373. |
Saile E, McGarvey J A, Schell M A, et al. 1997. Role of extracellular polysaccharides and endoglucanase in root invasion and colonization of tomato plants by Ralstonia solanacearum. Phytopathology, 87: 1264-1271. DOI:10.1094/PHYTO.1997.87.12.1264 |