 2007, Vol. 43
2007, Vol. 43文章信息
- 杨子祥, 陈晓鸣, 冯颖, 张燕平.
- Yang Zixiang, Chen Xiaoming, Feng Ying, Zhang Yanping.
- 倍蚜种间亲缘关系及角倍蚜种群分化的RAPD分析
- RAPD Analysis of Phylogenetic Relationships in Chinese Gallnut Aphids (Homoptera: Pemphigidae) and Genetic Differentiation in Four Populations of Schlechtendalia chinensis
- 林业科学, 2007, 43(7): 44-50.
- Scientia Silvae Sinicae, 2007, 43(7): 44-50.
-
文章历史
- 收稿日期:2005-11-21
-
作者相关文章



五倍子是倍蚜虫寄生在盐肤木(Rhus chinensis)、红麸杨(R. punjabensis var. sinica)和青麸杨(R. potaninii)等的树叶上,刺激叶组织细胞增生膨大而形成的虫瘿。倍蚜属同翅目(Homoptera),瘿绵蚜科(Pemphigidae),五节根蚜亚科(Fordinae);我国的倍蚜被划分为5属,14种(亚种)。其中角倍蚜(Schlechtendalia chinensis)是生产中利用最多的种。该蚜所形成的角倍,其产量约占我国五倍子总产量的75%(唐觉等,1957;向和,1980;张广学等,1999)。由于倍蚜体型微小,生活周期复杂,形态变异较大,其分类在学术界存在较大的争议(李治国等,2003)。RAPD技术能够简单、快速地检测生物间的遗传差异,其所需的DNA材料极少,对于研究蚜虫这样的微小昆虫十分有利(龚鹏等,2001),RAPD技术已被应用于蚜虫的种类鉴定、生物型识别和遗传变异研究(Black IV et al., 1992),但在倍蚜的研究方面,还没有相关的报道。
五倍子是生产可溶性单宁酸、没食子酸和焦性没食子酸等化工产品的重要原料,在医药、纺织、化工、食品、环保和农业等行业中用途广泛;我国五倍子产量约占世界总产量的95%,五倍子不仅是我国重要的资源昆虫产品和传统的出口商品,其寄主植物还有绿化荒山、保持水土的功效(赖永祺,1987)。因此,应用分子遗传标记技术,从DNA水平研究倍蚜的亲缘关系和种群分化,对于保护和合理利用这一生物资源具有重要意义。本研究采用RAPD技术,对11种倍蚜的亲缘关系和角倍蚜的4个地理种群间的差异进行了分析研究。
1 材料与方法 1.1 倍蚜样品采集与保存供试倍蚜样品采集于四川、陕西、贵州和云南(表 1);角倍蚜实验种群系春季从四川峨眉引进,在昆明温室接种到夏寄主盐肤木上,秋季倍子成熟后采集。为保证样品来自不同的克隆,在野外采集相隔1.5 m以上、接近成熟但尚未爆裂的倍子,带回室内打开,将孤雌胎生有翅或无翅成蚜转移到1.5 mL离心管中,无水乙醇浸泡,-20 ℃保存。采集的样本总数约200份,试验时随机抽取个体。
|
|

从11种倍蚜和角倍蚜4个地理种群样品中抽取116份样品,采用修改后的蛋白酶K法提取倍蚜的基因组DNA (田英芳等,1999;杨子祥等,2005;2006),测定DNA的浓度和纯度,采用琼脂糖电泳检测DNA的完整性,-20 ℃保存备用。
1.3 DNA扩增PCR反应体系为20 μL,其中含1×buffer,2.5 mmol·L-1 MgCl2,0.5 mmol·L-1 dNTP (promega),2U Taq酶(promega),66 ng引物(上海Sangon),1 μL模板DNA,加入ddH2O至20 μL。扩增在MJ-PTC 200 DNA扩增仪上进行,条件为:94 ℃ 5 min,后进行40个循环,即94 ℃ 1 min、36 ℃ 1 min、72 ℃ 2 min,最后于72 ℃延伸10 min,保存于4 ℃。在种间水平上,每个种扩增8~10头个体。在角倍蚜种群水平上,每个种群检测4个个体,用倍蛋蚜作外类群。
1.4 产物检测将扩增后的DNA在1.5%的琼脂糖凝胶上电泳,上样量为2.0 μL,电压为3 V·cm-1,时间为90 min,电泳缓冲液为1×TBE;0.05% EB染色,UVP GDS-8000凝胶成像系统观察、照相和分析。用GeneRuler 100 bp DNA Ladder Plus (Fermantas)作为分子量的标准。
1.5 数据统计与分析根据分子量大小对扩增结果读带,然后转变为可供计算机处理的0-1数据(分别表示特定位点上的有或无)。采用Popgene 1.31软件进行分析,计算遗传距离(D)和遗传一致性(I),遗传距离反映2个群体在趋异进化过程中发生等位基因替换位点的比例,它代表所有位点上基因分化水平的平均值,不同位点在群体间的遗传距离各不相同;遗传一致性是指2个群体间完全一致的基因所占的比例(郑哲民等,2000)。在Mega 3.1软件中用非加权成组配对法(UPGMA)对群体进行聚类分析。
2 结果与分析 2.1 PCR反应体系的建立为测试PCR反应体系的稳定性,先筛选出一条扩增条带清晰、多态性好的引物S7,对肚倍蚜指名亚种同一克隆4个个体的DNA进行扩增,用同一属的2个亚种和米倍蚜作对照。从图 1可以看出:4个个体的带型基本相同,分别在350、800、1 100和1 500 bp处各有4条清晰、明亮的条带。3个对照的带型是:肚倍蚜蛋铁亚种在350、1 100和1 500 bp与肚倍蚜指名亚种有共同的条带,但缺少800 bp的明亮条带;肚倍蚜蛋肚亚种在350和1 500 bp与肚倍蚜指名亚种有共同的条带,但缺少800、1 100 bp的条带;米倍蚜与3种肚倍蚜指名亚种之间只有350 bp的共同条带。这表明:只要引物合适,采用本PCR反应体系可以得到重复性和种间特异性较好的结果,以下的试验均采用此反应体系和条件。

|
图 1 随机引物S7扩增肚倍蚜同一克隆4个个体DNA的电泳图谱 Fig. 1 RAPD-PCR profiles obtained with primer S7 showing the variation among the genomic DNA of the four individuals M:标准分子量DNA Ladder.下同。The same below. I1-I4:肚倍蚜指名亚种K. rhusicola rhusicola (The same clone);O:米倍蚜M. microgallis;L:肚倍蚜蛋铁亚种K. rhusicola ovogallis;K:肚倍蚜蛋肚亚种K. rhusicola ovatirhusicola;CK:对照Negative control. |
从40条引物中筛选出13条条带清晰、多态性较好的引物进行扩增,共扩增出了200个位点,多态位点率为100%,平均每条引物扩增出15.4个位点,片段大小为100~3 000 bp(表 2)。表明11种倍蚜之间具有非常丰富的DNA系列多态性。
|
|
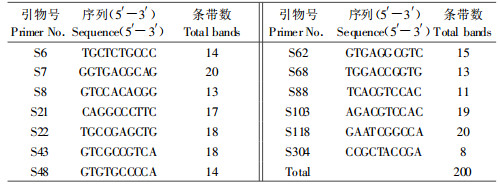
电泳图谱显示:属间带型差异明显,共有条带较少;属内一般具有相同的主带,共有条带较多。如引物S103的扩增结果是角倍蚜和倍蛋蚜在400和1 100 bp有共同的条带,与肚倍蚜枣铁亚种没有共有带(图 2a);引物S22的扩增结果是倍花蚜、红倍花蚜在400、550、900、1 200和1 400 bp有共同的条带,与肚倍蚜枣铁亚种仅在900和1 400 bp有共同的条带(图 2b),这2个条带可以看作这2个属间的共有条带;引物S62的扩增结果是黄小铁枣倍蚜和米倍蚜在400、800、1 400和1 800 bp有共同的条带,与红小铁枣倍蚜在400、800 bp处有共同的条带(图 2c),即本属的3个种有2条共有条带;引物S43的扩增结果是肚倍蚜指名亚种、肚倍蚜蛋肚亚种、肚倍蚜蛋铁亚种在600、900和1 300 bp有共同的条带(图 2d)。

|
图 2 不同引物扩增的DNA电泳图谱 Fig. 2 RAPD profiles obtained with various primer a:M:引物S103 Primer S103; E1-E10:角倍蚜S. chinensis; F1-F10:倍蛋蚜S. peitan; J1-J4:肚倍蚜枣铁亚种K. rhusicola ensigallis. b:M:引物S22 Primer S22; J1-J6:肚倍蚜枣铁亚种K. rhusicola ensigallis; G1-G9:倍花蚜N. shiraii; H1-H8:红倍花蚜N. yanoniella. c:M:引物S62 Primer S62; M1-M8:红小铁枣倍蚜M. elongallis; N1-N8:黄小铁枣倍蚜M. flavogallis; O1-O8:米倍蚜M. microgallis. d:M:引物S43 Primer S43. I1-I8:肚倍指名亚种K. rhusicola rhusicola; K1-K8:肚倍蚜蛋肚亚种K. rhusicola ovatirhusicola; L1-L8:肚倍蚜蛋铁亚种K. rhusicola ovogallis. |
从40条引物中筛选出9个条带清晰、多态性较好的引物进行扩增,共扩增出了133个位点,其中多态位点36个,多态位点率为27.7%,平均每条引物扩增出14.7个位点,片段大小为250~2 800 bp(表 3)。
|
|
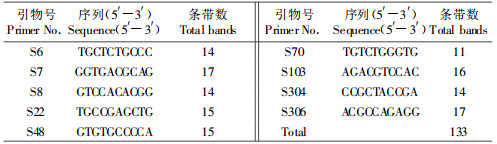
电泳图谱显示:角倍蚜4个地理种群具有非常相似的带型,它们与倍蛋蚜(外类群)的带型则明显不同。引物S48的扩增结果,4个地理种群在400、650和800 bp有共同的条带,仅在一些弱带上有差别(图 3);倍蛋蚜(外类群)虽在400、650和800 bp处与角倍蚜有共同的条带,但800 bp处的条带亮度较弱,且在1 031、1 200和1 500 bp还有3个条带。

|
图 3 随机引物S48扩增角倍蚜4个地理种群DNA的电泳结果 Fig. 3 RAPD profiles obtained with primer S48 showing the variation among the genomic DNA of the four geographic populations A1-A4:峨眉种群Emei, Sichuan; B1-B4:试验种群(昆明) Experimental population (Kunming); C1-C4:西乡种群Xixiang, Shaanxi; D1-D4:湄潭种群Meitan, Guizhou; F1-F4:倍蛋(外类群)S. peitan (Out-group). |
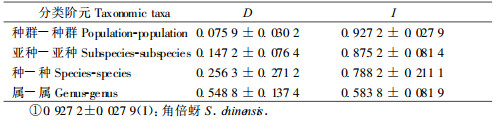
从表 4看出:倍蚜种间的遗传距离介于0.064 1~0.726 9之间,平均值为0.425 2,可见11个种之间存在较大的遗传差异;其中遗传距离在不同属之间为0.482 8±0.170 8,不同种之间为0.252 0±0.178 0,不同亚种之间为0.147 2±0.076 4,在角倍蚜种群间为0.075 9±0.030 2(表 5)。随着分类地位的接近,遗传距离逐渐减小;其中肚倍蚜的4个亚种具有很高的遗传一致性,其平均遗传距离为0.149 6,远远小于11个种间的平均遗传距离0.425 2,这与将它们调整为种下分类单元的结论相符合(张广学等,1999)。
|
|

|
|
从聚类图(图 4)可以看出:11种倍蚜聚集成2类,圆角倍蚜属的倍花蚜和红倍花蚜聚集成1类,其余9种倍蚜聚集成另一类,反映了它们在虫瘿类型上的差异。倍花蚜和红倍花蚜的虫瘿具有多重分枝,整个倍体呈花状,其他9种倍蚜的虫瘿为枣状、梨状或球状,暗示它们可能存在不同的起源;此结果与Zhang等(1999)认为圆角倍蚜属是首先分化的类群相符合。在另一分支中,肚倍蚜蛋肚亚种和肚倍蛋铁亚种首先聚在一起,后依次和肚倍蚜指名亚种、肚倍枣铁亚种聚成1类,显示了它们极为相近的亲缘关系;黄小铁枣倍蚜和米倍蚜聚成1支,然后和红小铁枣倍蚜聚成1类,显示了它们同为同一属的亲缘关系;角倍蚜与倍蛋蚜在较远的分枝上聚在一起,表明它们虽属于同一个属,但遗传差异较大;上述9种倍蚜在聚类图上最终相聚,暗示着它们可能存在共同的起源。总体上看:倍蚜同属的各个种都聚集在一起,聚类结果与形态分类结果基本一致,即本研究从DNA水平上为倍蚜的现行分类体系提供了新的证据。

|
图 4 11种倍蚜的UPGMA聚类树状图 Fig. 4 Dendrogram of UPGMA clustering based on genetic distances among 11 species of Chinese gallnut aphids |
分析表明:峨眉种群与试验种群的遗传距离最小(0.035 5),与陕西西乡种群和贵州湄潭种群的遗传距离依次增大(0.054 2和0.095 0)。这4个地理种群与外类群的遗传差异明显,其平均遗传距离为0.493 6(表 6)。Zhang等(2000)采用RAPD技术对蚜虫不同分类阶元的研究结果是种间的遗传距离为0.297 5 ± 0.062 7,同种的不同种群间为0.043 3 ±0.022 2。表 6说明角倍蚜4个种群间的遗传距离为0.075 9 ± 0.030 2,略大于同种的不同种群间的平均值,但小于种之间的平均值。表明角倍蚜的不同种群间存在着遗传分化,但其分化尚处于种群水平。
|
|

聚类图直观地显示了种群间的遗传差异和分化程度。峨眉种群与试验种群首先聚成一类,这与试验种群系从峨眉引种相符合,然后依次和陕西西乡种群和贵州湄潭种群相聚,最后才与外类群汇合(图 5)。可见角倍蚜的地理种群间已经出现了一定程度的遗传分化,造成这种分化的原因可能是地理上的隔离。陕西、四川、贵州和云南是我国倍蚜的起源和分布中心(张广学等,1999),其最初的分布可能是连续的,后来由于气候和寄主分布的变化,使得角倍蚜的分布不再连续,基因交流受阻,种群间产生了分化。四川与陕西接壤,其种群分化较晚,因而遗传差异较小,而贵州湄潭与陕西西乡距离较远,种群分化较早,其间还有长江的阻隔,因而遗传差异较大。

|
图 5 角倍蚜不同地理种群的UPGMA聚类树状图 Fig. 5 Dendrogram of UPGMA clustering based on genetic distances among 4 populations of Chinese gallnut aphids |
从RAPD研究结果看,倍蚜4个属间的差异明显(其平均遗传距离为0.482 8),这与形态分类的结果基本一致。但属下各种的亲缘关系的分子数据与形态分类并不完全一致,除倍蚜属外,另外3个属内的各个种间的遗传差异较小;圆角倍蚜属的倍花蚜与红倍花蚜的遗传距离为0.064 9,小铁枣蚜属的黄小铁枣倍蚜与米倍蚜的遗传距离分别为0.076 2,并在聚类图上先后聚成了1类,显示它们之间很近的亲缘关系和较高的遗传一致性,参照铁倍蚜属的分类模式和前人的研究结果(张广学等,1999;Zhang et al., 2000),将它们分别调整为种下分类单元似乎更为合理。
3.2 倍蚜对夏寄主树的选择与其亲缘关系没有明显的相关性在传统的倍蚜分类中,夏寄主的不同是倍蚜分类的依据之一,并把寄生于盐肤木上的虫瘿称为“角倍”,寄生于红麸杨和青麸杨上的分别称“铁倍”和“肚倍”(赖永祺,1987),但这种差异在RAPD分析时并不明显。黄小铁枣倍蚜和米倍蚜的夏寄主树分别为红麸杨和青麸杨,但在聚类图上聚成了1类,其遗传一致性为0.929 6,相似性较高,它们也许只是同1个种的2个寄主专化型。相反,角倍蚜和倍蛋蚜的夏寄主树都是盐肤木,却没有聚成1类,它们的遗传一致性只有0.638 2,可见对夏寄主树的选择与倍蚜种间亲缘关系没有明显相关性。
3.3 角倍蚜种群的分化较为明显角倍蚜4个不同地理种群间具有较高的DNA序列多态性和种群分化,其生物学和繁殖习性有助于保持种群间的遗传分化特性。在年生活史中需要冬、夏2类寄主,在夏寄主上形成五倍子;在其生活周期中,雌雄性蚜交配后,每头雌蚜只产1头干母,干母爬到夏寄主树上寄生形成虫瘿(倍子),并在虫瘿内营孤雌生殖(张广学等,1983),这种有性世代与无性世代,两性生殖与孤雌生殖交替的生活方式,使倍蚜的遗传特性不致于因为大量的孤雌生殖而丧失,保持了种群内遗传多样性的稳定性。
3.4 RAPD技术在倍蚜研究中的价值形态分类直观、简便,容易操作,仍是各种分类技术中的主导技术。但是,倍蚜微小、形态变异大,仅凭形态特征分类常有较大的困难。物种进化的本质是基因的进化,以分子技术来研究倍蚜的亲缘关系和分类比形态技术有优越性。RAPD技术具有很高的敏感性,能检测出倍蚜种群间的微小差异,因而非常适合于进行相似种的识别和种群分化的研究。但本研究仅仅考虑了扩增条带的位置和有无,忽略了其他一些信息;倍蚜分类只有综合应用形态学、细胞学和分子遗传标记技术,才能全面地反映其遗传变异和系统发育关系。
龚鹏, 杨效文, 谭声江, 等. 2001. 分子遗传标记技术及其在昆虫科学中的应用. 昆虫知识, 38(2): 86-91. DOI:10.3969/j.issn.0452-8255.2001.02.002 |
赖永祺. 1987. 五倍子丰产技术. 北京: 中国林业出版社, 1-65.
|
李治国, 杨文云, 夏定久. 2003. 中国五倍子研究现状. 林业科学研究, 16(6): 760-767. DOI:10.3321/j.issn:1001-1498.2003.06.017 |
唐觉, 蔡邦华. 1957. 贵州湄潭五倍子的研究. 昆虫学报, 7(1): 131-140. |
田英芳, 黄刚, 郑哲民, 等. 1999. 一种简易的昆虫基因组DNA提取方法. 陕西师范大学学报:自然科学版, 27(4): 82-84. |
向和. 1980. 中国青麸杨五倍子蚜虫的研究. 昆虫分类学报, 2(4): 303-313. |
杨子祥, 冯颖, 陈晓鸣. 2005. 一种有效的蚜虫基因组DNA提取方法. 林业科学研究, 18(5): 641-643. DOI:10.3321/j.issn:1001-1498.2005.05.025 |
杨子祥, 陈晓鸣, 冯颖, 等. 2006. 蚜虫基因组DNA提取方法的改进. 昆虫知识, 43(6): 880-884. DOI:10.3969/j.issn.0452-8255.2006.06.035 |
张广学, 乔格侠, 钟铁森, 等. 1999. 中国动物志:昆虫纲14卷.同翅目.纩蚜科.瘿绵蚜科. 北京: 科学出版社, 1-273.
|
张广学, 钟铁森. 1983. 中国经济昆虫志:第25册.同翅目.蚜虫类(一). 北京: 科学出版社, 1-80.
|
郑哲民, 黄刚. 2000. 螽NFDA2总科部份属种RAPD带型变异的比较研究. 陕西师范大学学报:自然科学版, 28(3): 96-99. |
Black IV W C, DuTeau N M, Puterka G J, et al. 1992. Use of the random amplified polymorphic DNA polymerase chain reaction (RAPD-PCR) to detect DNA polymorphisms in aphids. Bulletin Entomological Research, 82: 151-159. DOI:10.1017/S0007485300051671 |
Zhang Guangxue, Chen Xiaolin. 1999. Study on phylogeny of Pemphigidae (Homoptera: Aphidinea). Acta Entomologia Sinica, 42(2): 1-11. |
Zhang Sufang, Yang Xiaowen, Ma Jisheng. 2000. A study on genetic distance among different taxa of aphids (Homoptera: Aphidoidea). Entomologia Sinica, 7(3): 235-242. |