 2007, Vol. 43
2007, Vol. 43文章信息
- 汪贵斌, 曹福亮, 柳学军, 郝明灼, 胡光磊, 汪企明.
- Wang Guibin, Cao Fuliang, Liu Xuejun, Hao Mingzhuo, Hu Guanglei, Wang Qiming.
- 落羽杉种源木材微纤丝角和纤维形态的变异
- Microfibril Angles and Fiber Morphology Characteristics of Baldcypress Wood
- 林业科学, 2007, 43(6): 117-122.
- Scientia Silvae Sinicae, 2007, 43(6): 117-122.
-
文章历史
- 收稿日期:2005-09-29
-
作者相关文章


2. 国家林业局 北京 100174;
3. 江苏省林业科学研究院 南京 211153
2. State Forestry Administration Beijing 100174;
3. Jiangsu Academy of Forestry Nanjing 211153
微纤丝角为细胞次生壁S2层微纤丝排列方向与细胞主轴所形成的夹角(曹福亮,1997),或可表述为细胞壁中纤维素链的螺旋卷索与纤维轴之间的夹角(Walke et al., 1995),其角度愈小则细胞的抗张强度愈大。木材的微纤丝角对木材主要物理、力学性能和化学性质有着很大的影响(Fang et al., 2004)。微纤丝角是木材机械性能的主要决定因子之一, 特别是弹性模量和异向收缩性,微纤丝角与木材密度存在一定的相互关系,并与木材强度和硬度密切相关(Walke et al., 1995; Hirabawa et al., 1998)。单个管胞中微纤丝角与纸浆纤维的抗拉强度和伸缩性密切相关,微纤丝角小, 抗拉强度大,微纤丝角大,则伸缩性强(Walke et al., 1995; 洑香香等,2002)。纤维形态特征是研究木材材性的基础之一,也是木材利用的依据之一。纤维形态特征存在着较大的变异,不同树种、同一树种的不同个体间、种源间、无性系间及同株不同部位均存在着明显的差异(Panshin et al., 1980)。落羽杉(Taxodium distichum)是古老的孑遗植物,我国大约从1917年开始引种,落羽杉生长快、适应性强,适合我国广大地区作用材、防护及庭院观赏树种。目前,有关落羽杉方面的研究主要表现在落羽杉种源的种子特性、苗木生长和生物量差异、种源扦插生根能力变异、抗逆性、材积计算方法、密度效应规律、根膝的形成和功能及遗传性等方面(曹福亮等,1995;汪企明等,1993;1998;汪贵斌等,2002;2004a; 2004b; 2004c; 2004d)。而关于落羽杉不同种源间木材性质方面的研究尚未见报道。本文研究了16个落羽杉种源的木材微纤丝角和纤维形态的空间变异,从木材微纤丝角和纤维形态方面进行落羽杉早期的种源选择,为落羽杉工业用材林的发展提供理论依据。
1 材料与方法试验地设在江苏省东海县李埝林场,林场位于江苏省东海县西北部,黄淮海平原的东南缘,属中纬度暖温带湿润地区,林场土壤类型以棕壤为主,有效土层较浅(30~40 cm),土壤有机质含量4~6 mg·g-1,全N含量0.45~0.62 mg·g-1,速效P 5~29 μg·g-1,速效K低于50 μg·g-1。参试落羽杉有16个种源,种源编号分别为2、3、4、5、6、7、8、9、10、12、13、14、30、17、16、15。1992年育容器苗,1993年3月12—14日定植。采用完全随机区组设计,4个重复,株行距2 m×3.3 m。2004年9月28日进行每木检测,每个重复选标准木1株,进行树干解析、圆盘带回备用。
试材采集全过程均按GB 1927-1943-9进行。微纤丝角测定的木材来源于4株标准木,每一标准木取5个试样,总计20个样本。微纤丝角样片按年轮、分早晚材削制成1.5 cm×0.5 cm×0.1 cm㎝的小样片,采用(002)面X-射线衍射图法测定微纤丝角(阮锡根等,1982),测定用X-射线衍射仪型号为丹东产的DX-2000。采用0.6T法从(002)面衍射强度曲线上求微纤丝角(阮锡根等,1993)。
木材纤维测定的木材来源于4株标准木,每一标准木取5个试样。木材纤维形态的测定采用富兰克林离析法(Donaldson, 1992),把测定木材密度的样品削成火柴状的小木条,把小木条装入玻璃试管中,加入体积比为1:1的冰醋酸(CH2COOH)和双氧水(H2O2)混合液,在沸水浴中消煮30 min,直至单个纤维完全解离,将样品冲洗至中性,用番红试剂染色后,取少量具代表性的样品置于载玻片上,在尼康YS100数码生物显微镜上直接测定纤维宽度,在ZOOM645体视数码显微镜上直接测定其长度。每一试样随机测定100根纤维的长度和宽度,取平均值。
不同种源间木材微纤丝角、纤维形态差异的测定样品均取自不同种源胸径处(1.3 m)同一年轮(第12年)的木材。
2 结果与分析 2.1 落羽杉种源木材微纤丝角的变异及空间变化 2.1.1 不同种源落羽杉微纤丝角的变异从表 1可以看出,不同落羽杉种源木材微纤丝角大小不同。10号种源落羽杉在所有种源间木材微纤丝角最高,为35.8°,3号种源落羽杉在所有种源间微纤丝角最低,为29.9°,8、7、13、12、4和9号种源的微纤丝角分别超出最低的3号种源17.73%、12.04%、11.37%、10.37%、10.37%和10.37%。方差分析结果表明:不同落羽杉种源间木材微纤丝角存在极显著差异(F=7.98,Pr=0.000 1)。多重比较结果表明:10和8号种源与其他种源间木材微纤丝角的差异均达到了显著水平。变异系数分析结果表明:16个种源落羽杉微纤丝角的总变异系数为6.37%,总的变异幅度为28.5~36.6°,30号种源落羽杉微纤丝角的变异系数最大,为5.02%,8、13、15、5、6和12号的变异系数分别为4.85%、4.26%、3.37%、3.37%、3.35%和3.37%,17号的变异系数最小,为0.92%。
|
|

图 1为落羽杉木材微纤丝角纵向变异的测定结果(4个种源的平均值)。从图 1可以看出,落羽杉木材微纤丝角随着树干高度的变化而变化。在同一年轮内(第12年),随着树干高度的增加,微纤丝角逐渐减少。方差分析结果表明:落羽杉由基干到顶部不同树高木材的微纤丝角存在显著差异(F=4.38,Pr=0.016 8)。多重比较结果表明:从距树干0~3.5 m处的落羽杉木材微纤丝角变异不显著,到3.35 m以后才显著减小。

|
图 1 落羽杉木材微纤丝角的纵向变异 Fig. 1 Longitudinal variation of microfibril angle of baldcypress wood 图中字母表示落羽杉微纤丝角纵向变异的多重比较分析结果。 The letters are the result of multiple comparison of microfibril angle of baldcypress wood. |
图 2为落羽杉木材微纤丝角径向变异的测定结果(4个种源的平均值)。图 2表明:落羽杉胸径处木材微纤丝角也存在一定的差异。从树皮到髓心,落羽杉木材微纤丝角呈逐渐增加的趋势。方差分析结果表明:落羽杉胸径处不同部位的木材微纤丝角差异未达显著水平(F=1.64,Pr=0.22)。

|
图 2 落羽杉木材微纤丝角的径向变异 Fig. 2 Radial variation of microfibril angle of baldcypress wood 1~6分别表示从树皮到髓心的6个样点,分别表示第12、10、8、6、4和2 a的树。 1~6 denoted wood samples from bark to pith, and 1~6 expressed separately the wood of 12、10、8、6、4 and 2 a. |
不同种源落羽杉木材纤维长度、宽度和长宽比差异较大。16个种源落羽杉木材纤维长度的总平均值为2 184.83 μm,纤维宽度为39.53 μm,长宽比为5.68(表 2)。其中纤维长度最高的种源为2、30和4号,纤维宽度较高的种源为16、14和8号,而长宽比较大的种源为2、3和10号(表 2)。根据国际木材解剖学会的规定,木纤维长度小于900 μm为短纤维,900~1 600 μm为中级长度,大于1 600 μm为长纤维。因此,落羽杉木材纤维均属于长纤维。
|
|
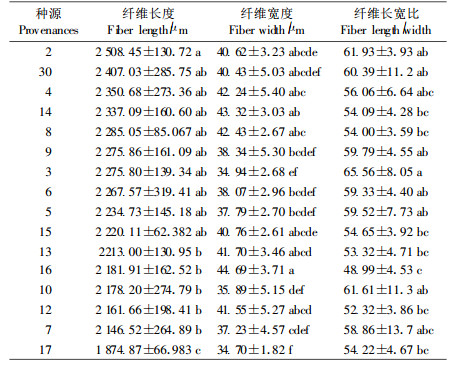
方差分析表明:16个种源落羽杉木材纤维长度、宽度和长宽比均达到显著差异水平(F=2.59,Pr=0.003 9;F=2.78,Pr=0.002 1;F=2.08,Pr=0.021 3)。
不同种源落羽杉木材纤维长度、宽度和长宽比等的变异系数差异也较大(表 3)。16个种源木材纤维长度、宽度和长宽比的总变异系数分别为13.04%、11.77%和13.98%,变异幅度分别为1 764.84~2 900.08 μm、28.48~51.21 μm和42.79~82.12(表 3)。6号种源木材纤维长度的变异系数最大,为14.09%,15号种源落羽杉木材纤维长度的变异系数最小,只有2.81%;木材纤维宽度的变异系数最大的种源为10号,为14.37%,最小的为17号种源,仅为5.27%;7号种源木材纤维长宽比的变异系数最大,比长宽比变异系数最小的2号种源高将近4倍。
|
|
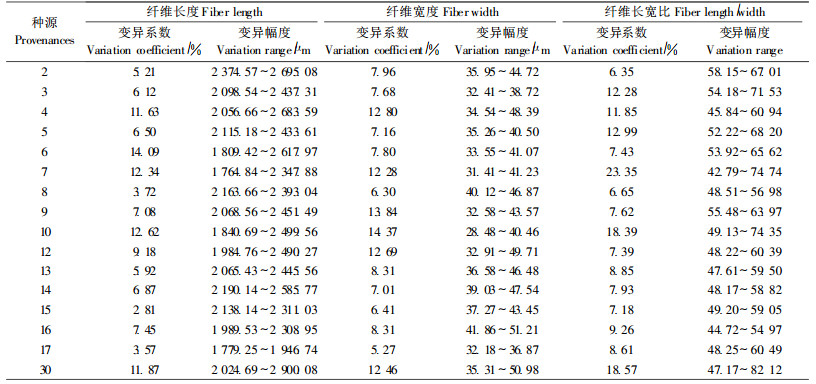
不同种源落羽杉木材纤维长度、宽度和长宽比等的变异幅度不同(表 3)。30号种源落羽杉木材纤维长度的变异幅度最大,为2 024.69~2 900.08 μm,17号种源木材纤维长度的变异幅度最小,为1 779.25~1 946.74 μm;30号种源落羽杉木材纤维宽度的变异幅度最大,为35.31~50.98 μm,17号种源落羽杉木材纤维宽度的变异幅度最小,为32.18~36.87 μm;30号种源落羽杉木材纤维长宽比的变异幅度最大,为47.17~82.12,8号种源落羽杉木材纤维长宽比的变异幅度最小,为48.51~56.98。
2.2.2 纤维形态的径向和纵向变异Pashin等(1980)在总结前人大量研究成果的基础上,将木材纤维和管胞径向变异归纳为3类:Ⅰ型,变化曲线是水平的,表示成熟细胞长度保持稳定不变;Ⅱ型,从幼龄材向成熟材过渡的曲线,表明长度逐渐增加;Ⅲ型,抛物曲线,表明细胞长度增加到最大值,然后减短。图 3显示了落羽杉胸径处木材的纤维长度由北到南的径向变异(4个种源的平均值)。从图中可以看出,胸径处纤维长度从髓心到树皮都表现为增加的趋势,径向变异规律应属PashinⅡ型,反映了树木生长发育过程中树龄对材性影响的一般规律,即在髓心附近,树木处于幼龄期,形成层原始细胞尚未成熟,细胞长度较短,形成层原始分裂较快,细胞长度逐渐增长,导致纤维长度迅速增长,年生长量也较大。

|
图 3 落羽杉木材纤维长度和宽度的径向变异 Fig. 3 Radial variation of fiber length and width of balcypress wood —○—纤维长度Fiber length, —△—纤维宽度Fiber width。下同The same below. 1~11表示胸径处从北到南不同样点,6为髓心,1~6分别表示第12、10、8、6、4和2 a的木材。1~11 denoted wood samples trom north to south at DBH, and 6 was pith, and 1~6 expressed separately the wood of 12、10、8、6、4 and 2 a. |
从植物生理学的角度看,纤维直径及胞腔径是和树木的生长发育相适应的。在树木生长的幼年期,树木内促进细胞分裂的激素多,顶端生长点活跃,纤维直径、腔径相对较大,随着树木生长发育的变化,激素也不断变化,到一定时期促进细胞延长的激素增多,细胞长度增长快,到后期抑制细胞增长的激素增多,细胞的增长受到抑制,细胞的增长呈稳定状态(王嘉楠,2002)。图 3显示了落羽杉胸径处木材的纤维宽度由北到南的径向变异。从图中可以看出,胸径处纤维宽度从髓心到树皮都表现为增加的趋势。
方差分析表明:胸径处纤维长度和纤维宽度的径向变异差异均达到显著水平(F=5.52,Pr=0.000 4;F=3.20,Pr=0.011 0)。
树木不同高度同一年轮内(第12年)纤维长度和宽度的测定结果(4个种源的平均值)见图 4。可以看出,纤维长度和宽度在同一年轮内不同高度间的变异均是随树高的增加纤维长度和宽度逐渐增加。

|
图 4 落羽杉木材纤维长度和宽度的纵向变异 Fig. 4 Longitudinal variation of fiber length and width of baldcypress wood |
相关分析结果表明:微纤丝角与木材纤维长度呈极显著的负相关关系(r=-0.4706,Pr=0.0075),与木材的纤维宽度和纤维长宽比呈一定的负相关(相关系数r分别为-0.032 9和-0.330 5),但未达到显著水平;木材纤维长度与木材纤维宽度呈显著的正相关(r=0.425 2,Pr=0.017 1)。
回归分析结果表明:微纤丝角(Y1)与木材纤维长度(X1)的线性回归方程为:Y1=44.3244-0.00533 X1 (F=8.25,Pr=0.0075),木材纤维长度(Y2)与木材纤维宽度(X2)的线性回归方程为:Y2=1502.3171+18.8082 X2 (F=6.4,Pr=0.017 1)。
3 结论与讨论对杉木(Cunninghamia lanceolata)、马尾松(Pinus massoniana)、湿地松(Pinus elliottii)等的研究表明:微纤丝角在髓心处最大,从髓心到树皮微纤丝角逐渐变小(鲍甫成等,1998)。另外,对杨树(Populus spp.)、银杏(Ginkgo biloba)、刺楸(Kalopanax septemlobus)、泡桐属(Paulownia)、红松(Pinus koraiensis)、黑杨(Populus nigra)、辐射松(Pinus radiata)、垂枝桦(Betula pendula)等的研究都表明微纤丝角在树木中的变化有以上相同的规律(郭德荣,1982; 刘盛全,1996; 刘盛全等,2001;孙成志等,1987;Fang et al., 2004; Donaldson, 1992; 1993; Bonham et al., 2001; 费本华等,2000)。许多研究也表明:微纤丝角在单株树木中的纵向变异是随树高的增加而逐渐下降的,但当达到一定的高度(不同树种有不同的高度)后微纤丝角基本达到一个较为稳定的值,但在树木梢头部微纤丝角又有所增大(洑香香等,2002)。本研究结果表明:16个种源落羽杉木材微纤丝角存在极显著差异,微纤丝角变化在29.9~35.8°之间,从径向变化看,落羽杉胸径处木材微纤丝角在髓心处(幼材处)最大,由髓心向外逐渐减小;从纵向变化看,微纤丝角在基干处最大,随树木高度的增加而逐渐减小。
木材纤维形态的变化受很多方面的影响,特别是树木生长发育的年龄阶段,纤维形态在树木幼龄阶段变化是非常活跃的,随着树龄的增大而迅速增加长度,到达一定年龄后达到稳定(朱教君等,1994;朱惠芳等,1962)。就造纸而言,纤维原料的纤维形态及其化学成分为纤维原料评价的重要依据。纸张强度与纤维形态有密切关系,过去认为纤维长度只要保持在330 μm以上,纸张的耐折度、撕裂指数、耐破指数等不再受其影响,但目前的研究表明:纤维长度越长,纸张的撕裂度、抗拉强度、耐破度和耐折度等也越强(朱教君等,1994)。本研究结果表明:16个种源的落羽杉木材的纤维长度、纤维宽度和纤维长宽比均存在极显著的差异,纤维长度的变化在1 764.84~2 900.08 μm之间,远远大于330 μm,从纤维长度来看,均是良好的造纸用材。纤维宽度的变化在28.48~51.21 μm,纤维长宽比的变化在42.79~82.12。国际木材解剖学规定,长宽比不低于35~45,能满足纤维工业原料的要求。因此,从长宽比来看,16个落羽杉种源基本上都能对满足纤维工业原料的要求。
16个落羽杉种源依据木材微纤丝角和木材纤维长度2个木材材性指标,进行系统聚类分析,从遗传距离0.9处可以分为4类:第Ⅰ类为10号和8号种源,木材的微纤丝角较大,木材纤维长度中等,作为纸浆工业用材林,不如其他种源;第Ⅱ类为12、7、13、4、9、14、15、16、5、6和3号种源,这11个种源的微纤丝角和木材纤维长度均处于中等水平;第Ⅲ类为2号和30号种源,这2个种源的木材微纤丝角较小,木材纤维长度比其他种源长,可作为纸浆工业用材林的优良种源;第Ⅳ类为17号种源,这个种源的木材纤维长度在所有种源中最短,微纤丝角中等。综合微纤丝角和木材纤维长度,2号和30号种源可作为纸浆工业用材林,特别是纸浆用材的优良种源。
鲍甫成, 江泽慧. 1998. 中国主要人工林树种木材性质. 北京: 中国林业出版社.
|
曹福亮, 刘成林. 1995. 美国落羽杉种源试验初报. 南京林业大学学报, 19(1): 65-70. |
曹福亮. 1997.林分密度对南方型杨树木材性质的影响//吕士行, 方升佐.杨树定向培育技术.北京: 中国林业出版社, 148-151
|
费本华, 江泽慧, 阮锡根. 2000. 银杏木材微纤丝角及其生长轮密度相关模型的建立. 木材工业, 14(3): 13-15. |
洑香香, 杨文忠, 方升佐. 2002. 木材微纤丝角研究的现状和发展趋势. 南京林业大学学报, 26(6): 83-87. DOI:10.3969/j.issn.1000-2006.2002.06.021 |
郭德荣. 1982. 红松微纤丝角变异与管胞长度和拉伸强度的关系. 东北林业大学学报, 10(2): 39-47. |
刘盛全. 1996. 刺楸木材微纤丝角与组织比量的变异研究. 安徽农业大学学报, 23(2): 186-190. |
刘盛全, 江泽慧, 鲍甫成. 2001. 人工林杨树木材性质与生长培育关系的研究. 林业科学, 37(2): 90-96. |
阮锡根, 尹思慈, 孙成志. 1982. 应用X射线衍射-(002)衍射弧法-测定木材纤维次生壁的微纤丝角. 林业科学, 18(1): 64-69. |
阮锡根, 王婉华, 潘彪. 1993. 应力木纤丝角的研究. 林业科学, 29(6): 534-536. |
孙成志, 谢国思. 1987. 泡桐属木材的微纤丝角和结晶度. 木材工业, 1(2): 24-28. |
汪贵斌, 曹福亮. 2002. 落羽杉抗性研究综述. 南京林业大学学报, 26(6): 78-82. DOI:10.3969/j.issn.1000-2006.2002.06.020 |
汪贵斌, 曹福亮. 2004a. 土壤盐分及水分含量对落羽杉生长的影响. 应用生态学报, 15(12): 2396-2400. |
汪贵斌, 曹福亮. 2004b. 盐分和水分胁迫对落羽杉幼苗生长量及营养元素含量的影响. 林业科学, 40(6): 56-62. |
汪贵斌, 曹福亮. 2004c. 土壤盐分及水分含量对落羽杉光合特性的影响. 南京林业大学学报, 28(3): 14-18. |
汪贵斌, 曹福亮. 2004d. 不同土壤水分含量下落羽杉根、茎和叶营养水平的差异. 林业科学研究, 17(2): 213-219. |
汪企明, 吕祥生. 1993. 落羽杉属种源研究:种子和苗期变异. 江苏林业科技, 20(1): 1-4. |
汪企明, 王伟. 1998. 落羽杉属种源研究:生长和生物量变异. 江苏林业科技, 25(1): 1-6. |
王嘉楠. 2002. 人工林杨树木材质量与生长培育关系的研究. 合肥: 安徽农业大学出版社.
|
朱蕙方, 李新时. 1962. 数种速生树种的木材纤维形态及其化学成分的研究. 林业科学, 7(4): 235-267. |
朱教君, 姜凤岐, 曾其蕴. 1994. 杨树林带木材纤维长度变化规律及其在经营中的应用. 林业科学, 30(1): 50-56. DOI:10.3321/j.issn:1001-7488.1994.01.017 |
Bonham V A, Barnatt J R. 2001. Fibre length and microfibril angle in silver birch. Holzforschung, (55): 159-162. |
Donaldson L A. 1992. Within and between tree variation in microfibril angle in Pinus radiada. New Zealand Journal of Forestry Sci, 22(1): 77-86. |
Donaldson L A. 1993. Variation in microfibril angle among three genetic group of Pinus radiata. New Zealand Jounal of Forestry Sci, 23(1): 90-99. |
Fang S Z, Yang W Z, Fu X X. 2004. Variation of microfibril angle and its correlation to wood properties in poplars. Journal of Forestry Research, 15(4): 261-267. DOI:10.1007/BF02844949 |
Hirabawa Y, Yamashita K, Fujisawa Y, et al. 1998.The effects of S2 microfibril angles and density on MOE in sugi tree logs//Butherfield B G.Microfibril angle in wood.New Zealand: Chritchurch, 312-322
|
Panshin A J, Carl de Zeeuw. 1980. Textbook of wood technology. 4th Edition. New York: Mc Graw Hill Book Company.
|
Walke J C F, Butterfield B G. 1995. The importance of microfibril angle for the processing industries. New Zealand Forestry, 40(4): 34-40. |