 2007, Vol. 43
2007, Vol. 43文章信息
- 伍维模, 李志军, 罗青红, 韩路.
- Wu Weimo, Li Zhijun, Luo Qinghong, Han Lu.
- 土壤水分胁迫对胡杨、灰叶胡杨光合作用-光响应特性的影响
- Effects of Soil Water Stress on Light Response Curves of Photosynthesis of Populus euphratica and Populus pruinosa
- 林业科学, 2007, 43(5): 30-35.
- Scientia Silvae Sinicae, 2007, 43(5): 30-35.
-
文章历史
- 收稿日期:2006-04-28
-
作者相关文章



2. 新疆生产建设兵团塔里木盆地生物资源保护与利用 重点实验室 阿拉尔 843300;
3. 塔里木大学植物科学技术学院 阿拉尔 843300;
4. 石河子大学生物工程学院 石河子 832003
2. Key Laboratory of Protection and Utilization of Biological Resources of Tarim Basin of Xinjiang Production & Construction Corps Alar 843300;
3. College of Plant Science and Technology, Tarim University Alar 843300;
4. College of Bioengineering, Shihezi University Shihezi 832003
胡杨(Populus euphratica)与灰叶胡杨(Populus pruinosa)为杨柳科(Salicaceae)杨属(Populus)的高大落叶乔木、稀灌木(王世绩,1996),是杨属中最古老、最原始的荒漠河岸林树种,为我国首批确定的388种珍稀濒危植物中的渐危种(苏培玺等,2003),具有防风固沙、稳定河道、保护绿洲、维持荒漠区脆弱环境生态平衡的功能。以胡杨、灰叶胡杨为主的河岸林是塔里木河干流两岸的主体森林类型,占全国胡杨林总面积89%,是我国胡杨林分布最集中的地区(王世绩,1996)。20世纪70年代以来,由于塔里木河流域大强度的水土资源开发利用,致使干流水量逐年减少,沿岸地下水位下降,土壤干旱加剧,引起塔里木河沿岸以地下水维系生存的天然胡杨林大面积衰败(陈亚宁等,2003)。因此保护与恢复塔里木河流域胡杨、灰叶胡杨林的关键措施是改善林地土壤水分状况。国内外对胡杨在渗透胁迫(马焕成等,1998a;Chen et al., 2002)、盐胁迫(Ma et al., 1997;马焕成等,1998b;Sixto et al., 2005)、自然生境条件下的光合作用及其光响应(Wang et al., 1997;马焕成等,1998c;Deng et al., 2003;Zhang et al., 2003)、水分生理特性(曾凡江等,2002;Gries et al., 2003)、对不同地下水位的生理反应(陈亚鹏等,2004)等方面做了较多的研究,但是有关胡杨、灰叶胡杨在水分胁迫下的光合作用-光响应特性研究还较少,尤其是灰叶胡杨的光合生理研究鲜见报道(Voznesenskii et al., 1981;刘建平等,2004;罗青红等,2006)。本文以2年生胡杨、灰叶胡杨幼苗为研究对象,探讨不同土壤含水量条件下胡杨、灰叶胡杨光合作用-光响应特性的差异,为林地土壤水分管理与天然林的更新复壮提供理论依据。
1 材料与方法 1.1 试验设计试验材料于2005年6月下旬采自新疆生产建设兵团农一师11团胡杨、灰叶胡杨苗圃。选择长势基本一致的2年生小苗,移栽于内径22 cm,高27 cm的生长盆中。每盆装相当于6.5 kg干土量的湿土,土壤采自苗圃,属于林灌草甸土,田间持水量26.47%。设3个土壤水分处理:正常供水(土壤相对含水量70%~80%,即土壤实际含水量占田间持水量的70%~80%)、中度水分胁迫(50%~60%)和重度水分胁迫(30%~40%)(表 1)。每一个土壤水分处理设置6个重复,胡杨、灰叶胡杨各栽植18盆,每盆栽植3株小苗,并在遮雨棚下培养,待苗木正常生长1个多月之后,开始按试验设计进行水分胁迫处理。由秤重法并结合取土烘干法来测定盆内土壤含水量。从8月6日开始,每天20:00用感量为千分之一的电子天平秤盆的质量,以此确定灌水量。盆内土壤含水量低于处理的下限含水量时,再补充水分,整个试验期间,盆内土壤的含水量始终控制在试验设计的范围之内。
|
|

水分胁迫处理10 d后,选择4个晴天上午在自然CO2浓度条件下(约375 μmol·mol-1),用LI-6400便携式光合作用系统(LI-COR,Inc,USA)的自动光曲线程序来测定各盆中上部叶片的光合作用-光响应。气体流速控制为0.5 L·min-1,叶室温度由LI-6400温度控制器控制在25±0.5 ℃,采用LI-6400红蓝LED作为光源。设定如下光合有效辐射(PAR):0、20、50、100、200、300、500、800、1 000、1 500、2 000、2 500、2 800、3 000 μmol photons·m-2s-1,测定叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)等光合生理生态参数,并计算水分利用效率(WUE)及羧化速率(CE)。
1.3 数据分析方法描述光合速率对光合有效辐射PAR响应曲线(即Pn-PAR曲线)的数学模型较多,如二次多项式模型(吴吉林等,2005)、指数模型(Bassman et al., 1991)、Michaelis-Menten方程(Mohr et al., 1995)、直角双曲线模型(王天铎,1990)和非直角双曲线模型(Farquhar et al., 1982)。经过对以上几种模型的比较,本文采用了非直角双曲线即Farquhar模型。
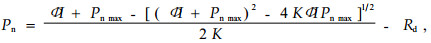
|
式中:Pn为净光合速率(net photosynthetic rate, μmol CO2·m-2s- 1),Pn max为一定CO2浓度下最大净光合速率(maximum photosynthetic rate, μmol CO2·m-2s-1);Φ为光响应曲线的初始斜率,即表观光合量子效率AQY(apparent quantum yield, μmol CO2·μmol-1 photons);I为光合有效辐射PAR(photosynthetically available radiation, μmol photons·m -2s-1);K为曲角,它描述了光响应曲线的弯曲程度;Rd为暗呼吸速率(dark respiration rate, μmol CO2·m-2s-1)。用SPSS统计软件中的非线性回归方法来估计模型参数(刘宇锋等,2005)。PAR在0~200 μmol photons·m -2s-1的Pn观察值近似一条直线,它与X轴(PAR)的交点就是光补偿点LCP(light compensation point, μmol photons·m-2s-1)(许大全,2002);该直线与Y=Pn max直线相交,交点所对应X轴的数值即光饱和点LSP(light saturation point, μmol photons·m-2s -1)(Walker,1989)。用SAS 6.12统计软件对模型参数进行方差分析,用Duncan氏新复极差法进行多重比较。
2 结果与分析 2.1 胡杨、灰叶胡杨光合作用-光响应特征参数由表 2可知正常供水条件下胡杨、灰叶胡杨的Pn max分别是24.5 9、16.68 μmol CO2·m-2s-1,明显高于中度和重度水分胁迫。胡杨与灰叶胡杨都生长在干旱荒漠地区,干旱和高光强相伴出现,即使是在3种不同的土壤水分条件下,胡杨Pn max始终比灰叶胡杨大,说明胡杨对高温、高光强及干旱的适应能力要大于灰叶胡杨。胡杨的光合能力强,Pn max较高,这有利于干物质的积累,为较高的物质产量的形成奠定了良好的生理基础。
|
|
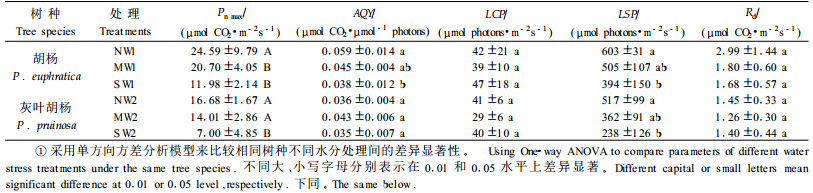
光合作用-光响应曲线最初部分的斜率即为表观光合量子效率AQY,它是光合作用中光能转化效率的指标之一(许大全,2002)。从表 2及图 1可知,在正常供水条件下胡杨的AQY值为0.059 μmol CO2·μmol-1 photons,该值明显地(P<0.05)高于灰叶胡杨(0.036 μmol CO2·μmol-1 photons)。水分胁迫使胡杨、灰叶胡杨AQY呈不同程度的下降。无论是正常供水还是水分亏缺,胡杨AQY均高于灰叶胡杨,说明胡杨对光能的转化效率高。光补偿点LCP是植物利用弱光能力大小的重要指标,该值越小表明利用弱光的能力越强。光饱和点LSP是植物利用强光能力大小的指标。具有高LSP与低LCP的植物,对光的生态适应能力强。在中度、重度水分胁迫条件下,与灰叶胡杨相比,胡杨均具有较高的LSP。暗呼吸速率Rd高低说明植物在没有光照条件下的呼吸速率不同(Coley,1983)。由表 3可知,水分胁迫对胡杨、灰叶胡杨的Rd无显著影响,但在相同土壤水分条件下,胡杨却显著(P<0.05)高于灰叶胡杨。胡杨与灰叶胡杨相比较,光合作用-光响应曲线的特征参数Pn m ax、LSP、AQY及Rd值均是胡杨显著地高于灰叶胡杨,而LCP二者之间无显著差异。水分胁迫极显著(P<0.01)或显著(P<0.05)降低了胡杨、灰叶胡杨的P n max、LSP、AQY,而对LCP和Rd无显著(P>0.05)影响。

|
图 1 水分胁迫对胡杨、灰叶胡杨表观光合量子效率(AQY)的影响 Fig. 1 Effects of water stress on apparent quantum yield(AQY) of P. euphratica and P. pruinosa Pn-PAR直线斜率即AQY。 The slope of Pn-PAR straight line represents AQY. |
|
|

从图 2a Pn-PAR响应曲线可见,控制在自然光强范围内,不同土壤水分条件下生长的胡杨、灰叶胡杨净光合速率Pn随着光合有效辐射PAR的增加呈上升趋势,在较低PAR(0~200 μmol photons·m-2s-1)时,增幅最大,当PAR达到一定的数值时,即到达两树种的光饱和点时,Pn曲线则渐趋平缓,出现光饱和现象。比较同一树种在不同水分处理之间Pn-PAR曲线的差异可知,胡杨、灰叶胡杨幼苗Pn值的大小均为正常供水>中度水分胁迫>重度水分胁迫,表明适宜土壤水分下生长的胡杨、灰叶胡杨幼苗叶片在弱光条件下对光强的变化敏感,随着测定光强的增大,正常供水下的幼苗叶片Pn仍然增加,而受水分胁迫的幼苗则在较低PAR时就达到了光饱和,Pn不再增加。在重度水分胁迫下胡杨、灰叶胡杨Pn-PAR响应曲线较为接近,走向较平缓。说明胡杨、灰叶胡杨幼苗的光能利用情况与土壤水分含量变化关系密切,干旱胁迫能降低幼苗的光能利用率。两树种叶片蒸腾速率Tr、气孔导度Gs、水分利用效率WUE、羧化速率CE均随PAR的增强而提高,这与光合速率随光强的变化一致(图 2b, c, e, f)。胞间CO2浓度Ci的变化趋势却与之不同,在PAR 0~500 μmol photons·m-2s-1时,随光强的增强迅速变小;在PA R 500~2 000 μmol photons·m-2s-1,Ci曲线的变化较为平缓;PAR高于2 000 μmol photons·m-2s-1后,Ci又表现出上升的趋势(图 2d)。这是由于Gs在前期提高的幅度较小,而光合速率不断加强使得Ci急剧降低,CO2供应赶不上光合速率增加的需求所致。随着光对气孔的刺激加强而导致气孔开张增大,Ci表现出上升趋势。随着PAR的不断增加,空气相对湿度降低,增大了叶片与大气间的水势差,使蒸腾速率增加,叶片内的水势降低。不同水分处理间以及相同水分条件不同树种间对光强的适应性亦不同,正常供水和中度胁迫下,两树种在较低光强刺激下,能够很快地使其气孔打开,蒸腾速率快速增加,从而使光合速率也迅速增加;但在重度水分胁迫下,气孔导度Gs随光强增大其变化较平缓。也就是说,当土壤严重干旱时,植物气孔基本上失去了调节作用,导致光合速率也出现对光强不敏感的现象。在相同土壤水分条件下,胡杨Gs、WUE、CE和Pn对光强的响应曲线始终高于灰叶胡杨;同一树种下,上述曲线的高低程度也基本上表现为正常供水>中度水分胁迫>重度水分胁迫。

|
图 2 不同水分处理下胡杨、灰叶胡杨净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)、水分利用效率(WUE)及羧化速率(CE)对不同光合有效辐射(PAR)的响应 Fig. 2 The light response curves of the net photosynthetic rate (Pn)、transpiration rate(Tr)、stomatal conductance(Gs)、intercellular CO2 concentration(Ci)、water use efficiency(WUE)、carboxylation efficiency(CE) to the photosynthetically available radiation (PAR)of the leaves of P. euphratica and P. pruinosa under water stress |
应用非直角双曲线模型(Farquhar模型)模拟了胡杨、灰叶胡杨在不同土壤含水量条件下光合作用-光响应曲线,并以此模型为基础计算了LSP、LCP、AQY、Pn max值。在正常供水条件下,胡杨、灰叶胡杨表观光合量子效率AQY分别为0.059、0.036 μmol CO2·μmol-1 photons,这一结果与丘国雄(1992)报道的C3植物取值范围(约0.055)基本接近。正常供水条件下胡杨、灰叶胡杨最大净光合速率Pn max分别为24.59、16.68 μmol CO2· m-2s-1,光饱和点LSP分别为603、517 μmol photons·m-2s-1,这一结果与Deng等(2003)在塔克拉玛干沙漠地区成年胡杨树的测定值基本一致。在正常水分条件下胡杨光饱和点LSP值却远远低于Ma等(1997)、马焕成等(1998c)报道的2 800 μmol photons·m-2s-1。其原因是:在拟合Pn-PAR曲线时,本文及Deng等(2003)所用的光响应模型是非直角双曲线模型,而Ma等(1997)、马焕成等(1998c)用的是一元二次回归模型,后者的LSP是曲线中Pn达到最大值时所对应的PAR值,从而使结果高于非直角双曲线模型值。
随着水分胁迫程度的加重,胡杨、灰叶胡杨的Pn max、AQY、LSP均不断下降,而LCP、Rd的变化不大。Pn max、AQY、LSP以正常供水最高,重度水分胁迫最低。表明水分胁迫使胡杨、灰叶胡杨叶的光合能力下降,对光强的利用范围变窄,降低了对光环境的适应能力。无论是在正常供水还是在水分胁迫下,胡杨AQY、Pn max、LSP、Rd高于灰叶胡杨,而LCP值无明显差异,表明胡杨比灰叶胡杨更能有效地利用弱光及强光进行光合作用,对光的生态适应性强,这是胡杨长期生存在干旱荒漠区对环境不断适应的结果。胡杨对于高光强的利用能力高于灰叶胡杨,在强光下,胡杨具有更高的光合速率与Pn max,光合同化潜力大。有研究表明,在一定的环境条件下叶片的Pn max表示了叶片的最大光合能力,而Pn max决定于Rubisco活性和电子传递速率(Walting et al., 2000)。在本试验条件下水分胁迫引起胡杨、灰叶胡杨Pn max、AQY下降,是否由于水分胁迫降低了PSⅡ电子传递速率及叶肉细胞Rubisco活性,还有待于深入的研究。
陈亚宁, 陈亚鹏, 李卫红, 等. 2003. 塔里木河下游胡杨脯氨酸累积对地下水位变化的响应. 科学通报, 48(9): 958-961. DOI:10.3321/j.issn:0023-074X.2003.09.017 |
陈亚鹏, 陈亚宁, 李卫红, 等. 2004. 塔里木河下游干旱胁迫下的胡杨生理特点分析. 西北植物学报, 24(10): 1943-1948. DOI:10.3321/j.issn:1000-4025.2004.10.032 |
刘建平, 韩路, 李志军, 等. 2004. 胡杨、灰叶胡杨光合、蒸腾作用比较研究. 塔里木农垦大学学报, 16(3): 1-6. |
刘宇锋, 萧浪涛, 童建华, 等. 2005. 非直线双曲线模型在光合光响应曲线数据分析中的应用. 中国生态农业学报, 21(8): 76-79. |
罗青红, 李志军, 伍维模, 等. 2006. 胡杨、灰叶胡杨光合及叶绿素荧光特性的比较研究. 西北植物学报, 26(5): 983-988. DOI:10.3321/j.issn:1000-4025.2006.05.018 |
马焕成, 王沙生. 1998a. 胡杨对渗透胁迫和盐分胁迫的不同响应. 西南林学院学报, 18(1): 1-7. |
马焕成, 王沙生, 蒋湘宁. 1998b. 盐胁迫下胡杨的光合和生长响应. 西南林学院学报, 18(1): 33-41. |
马焕成, 王沙生, 蒋湘宁. 1998c. 胡杨气体交换特性. 西南林学院学报, 18(1): 24-32. |
丘国雄. 1992. 植物光合作用效率//余叔文. 植物生理学和分子生物学. 1版. 北京: 科学出版社, 236-243
|
苏培玺, 张立新, 杜明武, 等. 2003. 胡杨不同叶形光合特性、水分利用效率及其对加富CO2的响应. 植物生态学报, 27(1): 34-40. DOI:10.3321/j.issn:1005-264X.2003.01.005 |
王世绩. 1996. 全球胡杨现状、保护与恢复. 世界林业研究, (6): 37-43. |
王天铎. 1990. 植物群落的光利用效率与数学模型//王天铎. 光合作用研究进展. 1版. 北京: 科学出版社, 128-211
|
吴吉林, 李永华, 叶庆生. 2005. 美丽异木棉光合特性的研究. 园艺学报, 32(6): 1061-1064. DOI:10.3321/j.issn:0513-353X.2005.06.018 |
许大全. 2002. 光合作用效率. 1版. 上海: 上海科学技术出版社, 33.
|
曾凡江, 张希明, Andrea Foetzki, 等. 2002. 新疆策勒绿洲胡杨水分生理特性研究. 干旱区研究, 19(2): 26-30. |
Bassman J B, Zwier J C. 1991. Gas exchange characteristics of Populus trichocarpa, Populus deltoides and Populus trichocarpa×P.deltoides clone. Tree Physiology, 11(8): 145-149. |
Chen Shaoliang, Li Jinke, Wang Tianhua, et al. 2002. Osmotic stress and ion-specific effects on xylem abscisic acid and the relevance to salinity tolerance in poplar. Journal of Plant Growth Regulation, 21(3): 224-233. DOI:10.1007/s00344-002-1001-4 |
Coley P D. 1983. Herbivory and defensive characteristics of tree species in a lowland tropical forest. Ecological Monographs, 53(2): 209-233. DOI:10.2307/1942495 |
Deng Xiong, Li Xiaoming, Zhang Ximing, et al. 2003. The studies about the photosynthetic response of the four desert plants. Acta Ecologica Sinica, 23(3): 598-605. |
Farquhar G D, Von C S. 1982. Modeling of photosynthetic response to environmental conditions//Lange O L, Nobel P S, Osmond C B, et al. Ecolopedia of plant physiology new series. Berlin: Springer-Verlag, 549-587
|
Gries D, Zeng F, Foetzki A, et al. 2003. Growth and water relations of Tamarix ramosissima and Populus euphratica on Taklamakan desert dunes in relation to depth to a permanent water table. Plant, Cell and Environment, 26(5): 725-736. DOI:10.1046/j.1365-3040.2003.01009.x |
Ma Huancheng, Lindsay F, Wang Shasheng, et al. 1997. Photosynthetic response of Populus euphratica to salt stress. Forestry Ecology Management, 93: 55-61. DOI:10.1016/S0378-1127(96)03943-6 |
Mohr H, Schopfer P. 1995. Plant physiology. 4th ed. Hong Kong: Springer-Verlag Hong Kong, 232.
|
Sixto H, Grau J M, Alba N, et al. 2005. Response to sodium chloride in different species and clones of genus Populus L. Forestry, 78(1): 93-104. DOI:10.1093/forestry/cpi009 |
Voznesenskii V L, Kremlev S M, Yudina O S. 1981. Characteristics of the carbon dioxide exchange of Kara-Kum plants. Soviet Plant Physiology, 28(2): 269-275. |
Walker D A. 1989. Automated measurement of leaf photosynthetic O2 evolution as a function of photon flux density. Philosophical Transactions of the Royal Society of London: Series B Biological Sciences, 323(1216): 313-325. DOI:10.1098/rstb.1989.0013 |
Walting J R, Press M C, Quick W P. 2000. Elevated CO2 induces biochemical and ultrastructural changes in leaves of the C4 cereal sorghum. Plant Physiology, 123(3): 1143-1152. DOI:10.1104/pp.123.3.1143 |
Wang Hongliang, Yang Shude, Zhang Chenglie. 1997. The photosynthetic characteristics of differently shaped leaves in Populus euphratica Olivier. Photosynthetica, 34(4): 545-553. DOI:10.1023/A:1006813513228 |
Zhang Lixin, Guo Jinkui, Li Wenrui, et al. 2003. Diurnal changes in photosynthetic characteristics of two differently shaped leaves in the desert plant Populus euphratica. Israel Journal of Plant Sciences, 51(4): 251-259. DOI:10.1560/MRB6-VV99-6DP9-Q4YA |