 2007, Vol. 43
2007, Vol. 43文章信息
- 刘丽华, 李保国, 齐国辉, 郭素萍, 赵志磊.
- Liu Lihua, Li Baoguo, Qi Guohui, Guo Suping, Zhao Zhilei.
- 雄性不育板栗雄花序败育与几种酶活性及MDA含量的关系
- Relationships Between Some Protective Enzymes Activities, MDA Content and Male Inflorescence of Male Sterile Chestnut
- 林业科学, 2007, 43(4): 121-124.
- Scientia Silvae Sinicae, 2007, 43(4): 121-124.
-
文章历史
- 收稿日期:2006-09-04
-
作者相关文章


2. 河北大学质量技术监督学院 保定 071002
2. College of Quality and Technology Supervision of Hebei University Baoding 071002
植物雄性不育广泛存在于开花植物有性繁殖过程中。据Kaul(1988)报道,已经在43科、162个属、320个种的617个品种或种间杂种中发现雄性不育。板栗(Castanea mollissima)也存在雄性不育现象。板栗既有雌雄混合花序,也有单纯的雄花序,并且雄花量极大,雌雄花序比例一般为1:1.7~12,而花朵比例则达1:2 400~4400(李保国,1998)。这些雄花的分化与发育,除消耗许多的糖分等有机营养外,还要消耗大量的氮、磷、铁及其他微量元素,严重影响雌花的分化与结实。而雄性不育可降低树体的营养消耗、省去去雄等繁琐手续等,在果树的生产和育种中具有十分重要的应用价值。因此,对板栗雄性不育机制的研究也就更为重要。
前人对果树雄性不育研究的报道不多,且大多集中于花粉母细胞减数分裂等细胞学过程,尚未见从生理生化方面研究木本植物雄性不育发生机制的报道。为了了解板栗雄性不育花序败育的生理机制,选用山东莒县林业局于2000年选育出的优良雄性不育板栗品种(王云尊等,2001)“浮来无花”和当地大量栽培的雄性可育品种“大红袍”,在雄花序发育的不同时期,测定了雄花序中SOD、POD、CAT活性及MDA含量的变化。
1 材料与方法 1.1 试验材料雄性不育板栗品种“浮来无花”和可育品种“大红袍”均为山东省日照市莒县小桑园村板栗园中的栽培品种,树龄为5年生。自雄花序出现(5月1日)开始取材,前期每隔3 d取一次样,3次以后每隔2 d取样一次,直至“浮来无花”雄花序脱落,两品种各取样8次。每次取样取雄花序约3 g,取下的样品立即放入冰壶,带回室内放入-20 ℃保存备用。
1.2 试验方法超氧化物歧化酶(SOD)活性测定采用NBT(氮兰四唑)光化还原法(李合生等,2003);过氧化物酶(POD)活性测定采用愈创木酚法(李合生等,2003);过氧化氢酶(CAT)活性测定用高锰酸钾滴定法(李合生等,2003);丙二醛(MDA)含量测定采用TBA(硫代巴比妥酸)法(邹琦,2001)。以各时期混合均匀的雄花序为测定材料,每样品重复3次。
2 结果与分析 2.1 雄花序败育过程中SOD活性的动态变化雄花序败育过程中SOD活性的变化如图 1所示。由图 1可以看出,两品种雄花序发育过程中SOD活性变化趋势基本一致,均为前期高后期低,且在同一时期,“浮来无花”雄花序的SOD活性均高于“大红袍”。“浮来无花”在5月18日时SOD活性最低,随后升高,到雄花序即将脱落的5月24日SOD活性又有所下降,为5月1日的93.1%。“大红袍”在5月15日时SOD活性最低,随后急剧升高,到5月21日又急剧下降。SOD是生物体内重要的保护酶之一,它的主要功能是催化超氧物阴离子自由基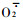
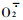

|
图 1 雄花序败育过程中SOD活性的变化 Fig. 1 Changes of SOD activities during male inflorescence development  浮来无花Fulaiwuhua, 浮来无花Fulaiwuhua,  大红袍Dahongpao.下同。The same below. 大红袍Dahongpao.下同。The same below.
|
雄花序败育过程中POD活性的变化如图 2所示。由图 2可知,“浮来无花”在雄花序败育过程中POD活性变化很大,在雄花序刚长出时POD活性比较高, 随后逐渐上升,至5月9日达到最高值14. 212 nmol·g-1FW,之后急剧下降,到5月15日POD活性最低,随后又急剧上升,到花序脱落时达到第二个高峰值13.181 nmol·g-1FW,仅次于5月9日。在雄花序发育过程中,“浮来无花”在8个取样时期中有6个时期POD活性高于“大红袍”。POD也是植物体内抗氧化保护酶系统的重要酶,SOD能清除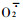

|
图 2 雄花序败育过程中POD活性的变化 Fig. 2 Changes of POD activities during male inflorescence development |
不育品种“浮来无花”在雄花序即将脱落时POD活性升高。POD活性在衰败过程中升高,曾有报道,月季切花衰老过程中POD活性是增加的(薛秋华等,1999),生姜叶片POD活性随叶龄的增大而升高(艾希珍等,1998)。但升高的原因及作用仍不很清楚。其原因可能有2个:一是POD利用SOD消除自由基时产生的H2O2,进行与衰老有关的氧化反应;一是POD与乙烯的自身催化合成有关,并且和衰老细胞的活性有关(莱谢姆等,1990)。

|
图 3 雄花序败育过程中CAT活性的变化 Fig. 3 Changes of CAT activities during male inflorescence development |
雄花序败育过程中CAT活性的变化如图 3所示。两品种雄花序发育过程中CAT活性变化趋势是一致的,且同一时期中“浮来无花”CAT活性明显高于“大红袍”。CAT也是植物体内消除自由基的酶类,SOD清除
雄花序败育过程中MDA含量的变化如图 4所示。由图 4可以看出,除雄花序即将脱落的5月24日外,“浮来无花”同一时期内MDA含量明显低于“大红袍”。因为“浮来无花”雄花序中发育前期SOD、POD、CAT活性高于“大红袍”,因而作为膜质过氧化产物的MDA含量也就相对较低(何宇炯等,1996)。随着“浮来无花”雄花序的发育,SOD、CAT活性降低,细胞内产生和清除自由基的能力失去平衡,活性氧含量增加,膜脂过氧化作用加强,MDA含量在雄花序即将脱落时达到最高,引起雄花序衰老、脱落,发生败育。而“大红袍”为雄性可育品种,雄花序MDA含量在5月24日时并未升高,反而有所下降。

|
图 4 雄花序败育过程中MDA含量的变化 Fig. 4 Changes of MDA contents during male inflorescence development |
1) 过去关于植物雄性不育的研究,主要集中在水稻(Oryza sativa)、高粱(Sorghum bicolor)、小麦(Triticum aestivum)等农作物以及一些草本植物方面,但是对于多年生木本植物雄性不育的研究相对较少,主要有杉木(Cunninghamia lanceolata)(吕洪飞等,1997)、桃(Prunus persica)(沈元月等,2002)、枣(Ziziphus jujuba)、芒果(Mangifera indica) (范眸天等,1996)、杏(Prunus armeniaca)(Lillecrapp et al., 1999)、橄榄(Canarium album)、柑橘(Citrus reticulata)(Yamamoto et al., 1995)等。国内关于板栗雄性不育的研究尚未见报道,国外报道也很少,其中Soylu (1992)的杂交结果认为板栗的雄性不育是由2个近乎等位的隐性基因控制的,并认为必须考虑2个基因的主效性关系和修饰基因的作用。种间杂种C.sativa(完全雄性不育)× C.crenata的后代只出现了无雄蕊型,不育类型没有出现分离现象(Bolvansky et al., 1999)。
有的研究认为,POD活性与雄性不育的败育有关。赵前程等(2002)对花椰菜雄性不育系进行了研究,结果认为不育花药的POD活性高于其保持系。认为过氧化物酶活性对控制体内的生长素(IAA)含量水平起着关键作用,而体内IAA含量水平直接关系到花药营养物质的积累和运输。因此不育系花药中过氧化物酶活性的增高,将导致花药内源激素(IAA)的氧化分解,从而影响营养物质的合成和运输,最终引起小孢子败育。此外,POD还与乙烯的自身催化合成有关,较高的POD活性可能促进了雄花序中乙烯的合成,进而导致雄花序的脱落。本试验中不育品种“浮来无花”雄花序败育过程中POD活性高于可育品种“大红袍”,与前人在农作物上的研究结果相似。但是SOD、CAT活性及MDA含量如何影响雄性不育进程还需进一步研究。
2) SOD、POD、CAT是植物中活性氧清除系统重要的酶,清除植物体内的自由基,维持自由基产生与清除系统的平衡。当这种平衡系统受到破坏时,细胞内出现自由基积累。大量积累的氧自由基以极其强烈的氧化作用诱发细胞膜中的不饱和脂肪酸,发生过氧化作用,其产物是具有强氧化性的脂质过氧化物和各种小分子的降解产物,尤其是丙二醛(MDA)含量增高最为显著。最终组织细胞膜完整性因之解体、破坏,细胞组分出现大量外渗,代谢紊乱至细胞伤害而死亡。在本研究中,“浮来无花”雄花序生长初期,SOD、POD、CAT活性都较高,随着花序的生长活性逐渐下降,到雄花序脱落时,SOD、CAT活性下降到最低,POD活性有所上升,而丙二醛含量则达到最高。这种活性氧清除系统的破坏可能是导致“浮来无花”雄花序萎蔫、脱落,最终发生不育的生理原因。对于这些物质的详细变化情况有待于深入研究。
艾希珍, 张振贤, 崔志峰, 等. 1998. 生姜叶片发育过程中光合速率与MDA含量及保护酶活性的变化. 园艺学报, 25(3): 294-296. |
范眸天, 李树云, 李云. 1996. 芒果雄性不育现象及花粉萌发研究初报. 云南农业大学学报, 11(1): 25-27. |
何宇炯, 徐如涓, 赵毓橘.1996.植物生理学报, 22(1): 58-62 http://www.cqvip.com/Main/Detail.aspx?id=2195685
|
莱谢姆Y Y, 哈勒维A H, 法伦克尔C.1990.植物衰老过程和调控.胡文玉, 译.沈阳: 辽宁科学技术出版社
|
李保国. 1998. 太行山板栗集约栽培. 北京: 中国林业出版社.
|
李合生, 孙群, 赵世杰, 等. 2003. 植物生理生化实验原理和技术. 北京: 高等教育出版社.
|
吕洪飞, 余象煜, 李平. 1997. 杉木雄性不育株与可育株小孢子囊发育的电镜研究. 武汉植物学研究, 15(2): 97-102. |
沈元月, 王东昌. 2002. "川中岛"白桃雄性不育小孢子发育的细胞形态学. 应用与环境生物学报, 8(4): 435-437. |
王云尊, 马元考, 陈维峰. 2001. 珍稀板栗新品种"浮来无花"的性状及栽培技术. 林业科技开发, 15(5): 31-32. |
薛秋华, 林如. 1999. 月季切花衰老与含水量、膜脂过氧化及保护酶活性的关系. 福建农业大学学报, 28(3): 304-308. |
赵前程, 耿宵, 陈雪平, 等. 2002. 花椰菜雄性不育系小袍子发育过程及其POD活性. 华北农学报, 17(2): 108-111. |
邹琦. 2001. 植物生理学实验指导. 北京: 中国农业出版社.
|
Bolvansky M, Mendel L. 1999. Inheritance of some fruit and flower chatacteristics in full sib progenies of chestnut. Acta Hort, 494: 344-399. |
Kaul M L H. 1988. Male sterility in higher plants. Berling: Springer-verlag Press.
|
Lillecrapp A M, Wallwork M A, Sedgley M. 1999. Female and male sterility cause low fruit set in a clone of the 'Trevatt' variety of apricot (Prunus aremeui). Sci Hort, 82(3/4): 255-263. |
Soylu A. 1992. Heredity of male steriliy in some chestnut cultivars. Acta Hort, 317: 181-185. |
Yamamoto M, Matsumoto R, Yamada Y. 1995. Relationship between sterility and seedlessness in Citrus. J Japan Soc Hort Sci, 64(l): 23-29. |