 2007, Vol. 43
2007, Vol. 43文章信息
- 何小勇, 柳新红, 袁德义, 谭晓风, 赵思东, 张琳, 魏来.
- He Xiaoyong, Liu Xinhong, Yuan Deyi, Tan Xiaofeng, Zhao Sidong, Zhang Lin, Wei Lai.
- 不同种源翅荚木的抗寒性
- Cold Resistance of Zenia insignis from Different Provenances
- 林业科学, 2007, 43(4): 24-30.
- Scientia Silvae Sinicae, 2007, 43(4): 24-30.
-
文章历史
- 收稿日期:2007-01-09
-
作者相关文章



2. 浙江省丽水市科普工作指导站 丽水 323000;
3. 浙江省林业科学研究院 杭州 310023
2. Lishui Popular Science Station of Zhejiang Lishui 323000;
3. Zhejing Forestry Academy Hangzhou 310023
翅荚木(Zenia insignis)又名任豆树、任木、砍头树,属苏木科(Caesalpiniaceae)翅荚木属落叶高大乔木,是我国南方可发掘利用的一个珍稀树种,为国家二级保护野生植物,自然分布地为桂、粤、湘、黔、滇,属于北热带和南、中亚热带地区(陈永密等,1989)。翅荚木具树干通直、生长迅速、萌芽性强、天然更新好、抗性强、耐瘠薄等诸多优良特性,被各地广泛引种栽培(覃志刚等,1991;何义发等,1996;曾广腾等,2004),成为当地速生丰产林及短轮伐期工业原料林建设的理想选择树种(余光等,1998;侯伦灯等,2001;周志春等,2003)。由于翅荚木自然分布于北热带和南、中亚热带地区,在江西、湖南、福建、浙江等地引种后遇冬季低温时易受冻害,特别是苗期和幼林期尤为严重,但是不同种源之间的冻害差异极大(陈亮明等,1997;柳新红等,2005)。因此,不同种源的抗寒性研究已成为翅荚木引种造林的重要内容。
目前,评价林木抗寒性的指标包括形态结构、生理代谢、抗寒基因等诸多方面(王利等,2002),而测定抗寒性的方法主要有全株冰冻测试法、电解质渗出率法、叶绿素荧光法、热分析法等(张钢,2005)。田间测定是评价林木越冬抗寒性最直观可靠的方法,冻害指数是常用指标。电导率的变化反映出质膜伤害的程度和所测材料抗寒性的大小,在油松(Pinus tabulaeformis)(唐季林等,1989)、马尾松(P. massoniana)(荣文琛等,1999)、印楝(Azadirachta indica)(廖声熙等,2004)等林木种源抗寒性评价中得到应用。有关植物叶片的组织结构与抗寒性之间的关系已经在其他林木上有过研究。这些研究大部分显示叶片细胞结构紧密度(cell tense ratio,CTR)和疏松度(spongy ratio,SR)、气孔密度等与植株的抗寒性有一定的关系(黄以江等,1982;简令成,1986;吴国良等,1998;吴林等,2005),但也有少数研究认为没有明显的相关性(刘世彪等,2004)。翅荚木抗寒性的研究,仅见陈亮明等(1997)进行了2个种源的田间耐寒力试验,肖双燕等(1992)初步分析了树高、地径、冠幅与冻害关系,有关翅荚木的叶片结构、叶片生理指标及它们与抗寒性的关系均未见报道。本文对国内5个种源的翅荚木的叶片进行了解剖研究,并测定了其田间抗寒性和叶片半致死温度、过冷温度,探讨了翅荚木的叶片结构特征与抗寒性的相关性,旨在为翅荚木抗寒种源的选择提供相应的依据。
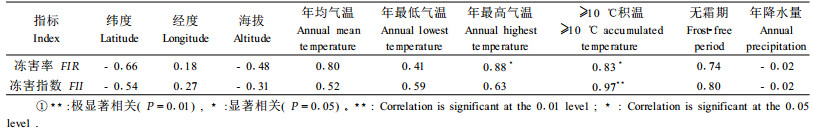
1 材料与方法 1.1 试验材料和试验设计翅荚木5个种源,分别于2004年9—10月采集于湖南省通道、广东省翁源、广西壮族自治区的平果、忻城和桂林。不同种源的地理位置和气候因子见表 1。
|
|
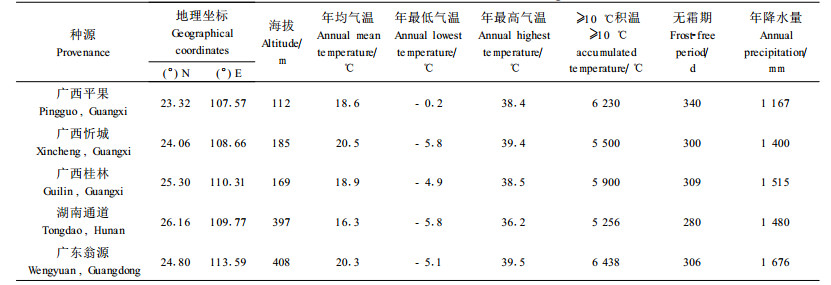
种源育苗试验点分别设置在中南林业科技大学湖南省株洲试验基地和浙江省丽水市林业科学研究所苗木基地。株洲位于113.16° E,27.93° N,属亚热带季风性湿润气候,历年均温17.5 ℃,极端最高温39.5 ℃,极端最低温-8.0 ℃,≥10 ℃年积温5 300 ℃,无霜期286 d,年降水量1 392 mm。株洲试验地海拔70 m,为缓坡丘陵红壤。丽水位于119.68° E,28.30° N,属中亚热带山地暖湿季风气候,历年均温18.1 ℃,极端最高温41.5 ℃,极端最低温-7.7 ℃,≥10 ℃年积温5 727 ℃,无霜期237 d,年降水量1 379 mm。丽水试验地海拔71 m,为沙壤质水稻土,较肥沃。试验苗木于2005年4月中旬播种,播种前1 d将供试种子用80~90 ℃热水浸种,自然冷却后继续浸泡24 h。浸种后吸胀的种子进行筑畦撒播,覆土0.5 cm,覆盖芒萁叶保湿。种子发芽幼苗出现第1对真叶时进行移植,株距20 cm,行距30 cm。苗木生长期间施肥2次(用尿素150 kg·hm-2),锄草2次,防治蚜虫1次。株洲试验点分种源随机排列;丽水试验点按完全随机区组设计,每小区10行,5次重复。
1.2 田间抗寒测定2006年3月,分别在两试验点调查苗木的冻害情况。株洲试验点参照陈亮明等(1997)的翅荚木受冻害等级划分标准进行调查。0级:未受冻或梢略受冻,冻害级值(freezing injury level,FIL)为0;Ⅰ级:地上部分冻死不足1/2,冻害级值为1;Ⅱ级:地上部分冻死达1/2以上,但未全部冻死,冻害级值为2;Ⅲ级:地上部分全部冻死,但根部未冻死,冻害级值为3;Ⅳ级:整株冻死,冻害级值为4。株洲试验点调查时分种源统计植株不同受冻级别的株数,计算出平均冻害级值和冻害指数(freezing injury index,FII)。平均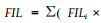
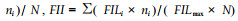
低温诱导:试验通过测定人工气候箱培养的经低温诱导的翅荚木小苗叶片的电导率来推测半致死温度。2006年9月中旬,将忻城、通道、翁源3个种源的种子浸种后播种于珍珠岩基质的容器中,置于人工气候箱内培养。根据翅荚木喜温耐热的特性,保持温度30 ℃,湿度90%,昼夜全光照强度为3 400 lx。真叶展开后,每周用Hoagland营养液浇1次。10月下旬,按每天5 ℃梯度降温至15 ℃时保持7 d,然后按每天2 ℃的梯度降至4 ℃时再保持7 d,进行低温诱导。
相对电导率测定:参考有关文献(王晶英等,2003),每种源取低温诱导后植株5株,各取叶片若干冼净擦干,随机分5组装入密封的塑料袋中。将其中1组置于4 ℃培养箱中作对照;其余4组分别置于0 ℃、-4 ℃、-8 ℃、-12 ℃进行低温处理24 h,处理结束后取出的样品放置于4 ℃的冰箱内解冻12 h,用直径为0.6 cm左右的打孔器打成小圆片。每种源各取小圆片10片放入25 mL的三角瓶中,加入无离子水20 mL,密闭,抽真空10 min,置于25 ℃水浴锅内平衡2 h,用EC200型电导/TDS/盐度测定仪测定其电导值C1,再将其煮沸30 min,冷却至25 ℃,测定其电导值C2。相对电导率=(C1-C)/(C2-C)×100%,式中:C为无离子水的导电值。试验重复3次,按种源分别计算不同温度处理下的平均相对电导率。
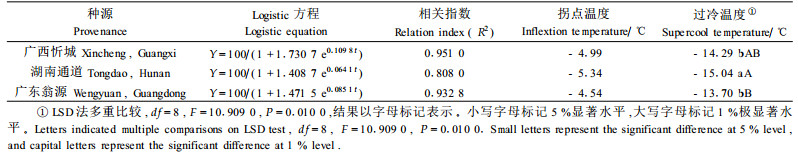
半致死温度计算:按缴丽莉等(2006)的方法,分种源将不同温度处理下的平均相对电导率拟合成Logistic方程Y=K/(1+ae-bt),式中:Y为低温处理下的相对电导率(%),t为处理温度(℃),K、a、b为参数。设定K值为100。求该方程的二阶导数并令其为0,则可获得曲线拐点(t,50),此时t即为拐点温度,与半致死温度(semi-lethal temperature,LT50)相一致,t=ln(a)/b。
过冷温度的测定:每种源取低温诱导后植株5株,分别取中部叶片上的小叶1张冼净擦干,重叠后用0.4 cm的打孔器取出小圆片,放入DSC Q10 V8.1型差示热量扫描仪测量铝皿内,以空铝皿为参比物,以1 ℃·min-1速度降温,测定其热流曲线,获得其相变时的放热峰,为叶片组织的过冷点温度。重复3次,采用最小显著性差异法(LSD)进行比较分析。
1.4 叶片解剖采样和制片分析叶片采样:2005年8月上旬从中南林业科技大学苗圃的1年生翅荚木苗木上采集叶片样本。每种源随机选取5株,每株统一选取离地高50 cm处的发育成熟的羽叶,摘取中部的小叶1张作样本。
石蜡制片及叶片解剖结构观察:每叶从小叶的中部切取1 cm×0.5 cm,用FAA固定液固定,常规石蜡制片法切片。切片厚度12 μm,番红、固绿双重染色,中性胶固封。制片后,每叶取一张代表性切片在Motic DMB5型数码生物显微镜下观察照相,用数码测距软件Motic Images Advanced 3.0测量叶片表皮层、栅栏组织和海绵组织的厚度,每项指标重复测定10次,取平均值。然后,计算每叶的细胞结构紧密度CTR和疏松度SR。CTR=(栅栏组织厚度+下部紧密组织厚度)/叶片厚度×100%,SR=海绵组织厚度/叶片厚度×100%。
叶表皮制片及气孔、表皮毛观测:每叶从小叶中脉一侧切取1 cm×0.5 cm,放入10%硝酸+10%三氧化铬混合溶液(40~50 ℃)充分氧化,离析表皮,修片后0.5%番红水溶液染色,二甲苯脱水,中性胶固封。每片选取10个视野显微观测气孔大小和密度,测量表皮毛长度和数量,计算每叶的平均值。
所有的解剖观测数据采用SPSS进行统计分析,以单株为小区,种源为因素,按随机区组设计进行统计分析,其中CTR和SR数据经反正弦平方根转换。
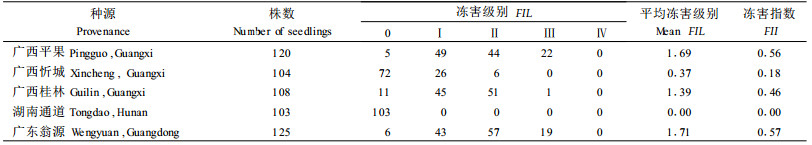
2 结果与分析 2.1 不同种源翅荚木的田间抗寒性比较翅荚木在湖南株洲播种育苗后,苗木生长良好,冬季低温对部分种源造成了一定的影响,不同种源抗寒性差异明显(表 2)。湖南通道种源无冻害;广西忻城种源冻害较轻,平均冻害级别为0.37,冻害指数为0.18;而其他种源平均冻害级别均>1,以广东翁源种源受冻最为严重,平均冻害级别为1.71,冻害指数为0.57。
|
|
翅荚木在浙江丽水播种育苗后,前期生长良好,12月份遇连续低温大部分种源苗木冻害严重。方差分析表明不同种源的冻害率差异达到了极显著水平。其中,湖南通道种源的冻害率平均为4.83%,极显著低于其他种源;广西忻城种源冻害率为73.61%,极显著低于广东翁源、广西平果和桂林种源;而广东翁源、广西平果和桂林种源间差异不大;冻害率最高的为广东翁源种源,达到了97.52%(图 1)。

|
图 1 不同种源翅荚木苗木冻害率比较 Fig. 1 Cold injury percentage of Z. insignis seedlings from different provenances 1.广西平果Pingguo, Guangxi; 2.广西忻城Xincheng, Guangxi; 3.广西桂林Guilin, Guangxi; 4.湖南通道Tongdao, Hunan; 5.广东翁源Wengyuan, Guangdong. |
由于2个试验点的气候条件不一样,同一种源的冻害情况有较大的差异。2005年12月中旬,丽水连续7 d的日最低气温均低于0 ℃,最低气温达-6.1 ℃,而同年株洲没有出现极端天气。在丽水播种育苗的翅荚木苗木冻害普遍比在株洲冻害严重。如湖南通道种源的翅荚木在株洲无冻害,而在丽水就有极少量的苗木受冻。因此,在浙西南山区种植翅荚木,需要选择耐寒种源进行引种试验。综合2个试验点的苗木抗寒性表现,可将5个翅荚木种源分为3大类:第1类是抗寒种源,采集于中亚热带湖南通道,其种源的耐寒力极显著强于其他种源,这主要是自然选择的结果;第2类是抗寒性一般的种源,采集于广西忻城;第3类是抗寒能力较差的种源,包括广西平果、桂林和广东翁源种源,苗期的田间抗寒性差。相关性分析(表 3)表明:不同种源的冻害指数与≥10 ℃有效积温极显著相关,冻害率与年最高气温、≥10 ℃有效积温显著相关,而与其他地理位置和气候因子没有显著相关性。
|
|
试验表明:翅荚木不同种源叶片组织经一系列低温处理后,相对电导率变化趋势基本一致。随着处理温度的下降,细胞内相对电导率上升,相对电导率的上升是零下低温对叶片原生质膜透性破坏的效应。低温处理初期相对电导率上升缓慢,然后在一定低温范围内相对电导率急剧升高,随后又趋于缓慢,即相对电导率随着温度的下降呈“S”型曲线变化。对叶片组织在不同低温处理下的相对电导率随温度变化用Logistic方程拟合,并求出其拐点(表 4)。一般认为,拐点温度值越低,表明其半致死温度越低,即该种源抗寒性越强。从表 4可以看出,湖南通道种源的拐点温度为-5.34 ℃,低于其他种源0.35~0.80 ℃,因此,湖南通道种源比其他种源具有较好的抗寒性。
|
|
利用植物细胞液在低温下结成冰晶伴随的放热现象测定植物组织的抗寒性,近20年来在国外已经陆续有相关的试验研究(Ashworth et al., 1981;Warmund et al., 1990;Kang et al., 1998;张善江等,2005),并出现了计算机差异热分析系统,国内也有学者采用自制的试验装置观测植物的超冷现象(余观夏等,2006)。一般认为植物组织的过冷点越低,植物抗寒性越强。试验表明,翅荚木不同种源叶片组织的过冷点不同。湖南通道种源叶片组织的过冷点为-15.04 ℃,低于其他种源0.75~1.34 ℃,差异达到显著水平,因此,湖南通道种源比其他种源具有较强的抗寒性。
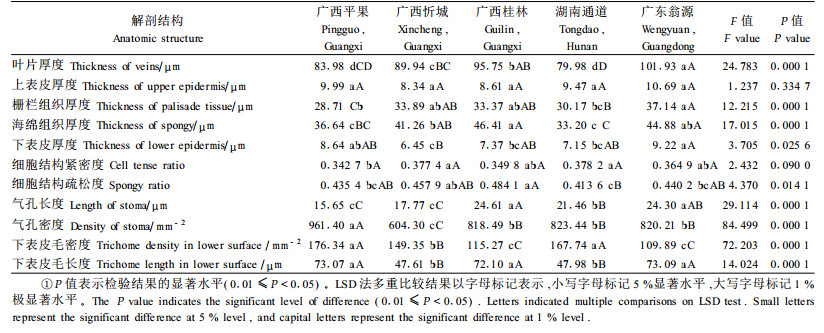
2.3 翅荚木的叶片解剖特征及其不同种源间的差异石蜡切片观察发现:不同种源翅荚木叶片的横切面基本结构是一致的。表现为主脉由角质层、表皮细胞、薄壁组织及维管组织组成。维管组织发达,由维管束鞘和维管束组成,排列成半圆形,维管束的韧皮部和木质部分化明显。叶片为异面叶,表皮由单层细胞构成。叶肉组织中的栅栏组织由1层细胞构成,呈圆柱形,排列紧密,约占叶肉的1/3~1/2;海绵组织细胞较大,不规则排列,细胞间隙发达,约占叶肉的1/2~2/3。
表皮制片观察发现:不同种源翅荚木的叶片表皮细胞形态特征基本相似。表现在表皮细胞1层,多为矩形、多角形,直径15~25 μm,无规则排列;细胞垂周壁呈U形、V形起伏;细胞表面纹饰局部加厚,上表皮比下表皮明显。表皮上分布有气孔,呈椭圆形,由2个半圆形的开口、保卫细胞组成,主要集中在下表皮,在上表皮上仅有零星分布。表皮上还有表皮毛,表皮毛无头部,只有柄部和根部,柄部为单细胞,根部为圆钉状,基部细胞常特化为辐射状,在上下表皮均有分布;上表皮大部分为单细胞的先端钩状的短表皮毛,夹生少量的长毛;下表皮毛由少量的短钩状毛和略弯曲或直立的长毛组成。
方差分析表明:不同种源的叶片厚度、栅栏组织厚度、海绵组织厚度、气孔长度、气孔密度、下表皮毛密度和下表皮毛长度存在极显著差异;CTR、SR和下表皮层厚度有显著差异;上表皮层厚度差异不显著(表 5)。从表 5可以看出:广东翁源种源叶片大而厚,达101.93 μm,相应的栅栏组织、海绵组织也比较厚,气孔也比较长,可达24.30 μm,表皮毛也比较长,达73.09 μm,但其CTR、SR值一般,表皮毛密度小,为109.89根·mm-2。湖南通道种源叶片较小而薄,平均叶片厚度为79.98 μm,只有其他种源叶片厚的78.47%~95.25%,但其CTR值大,达0.378 2,是所有种源中最大的;SR值小,只有0.413 6,是所有种源中最小的。说明通道种源叶片栅栏组织所占的比例较大,叶片结构紧密。同时,从表 5中还可以看出,湖南通道种源的表皮毛密度也比较大。相关分析表明:试验观测的叶片主要解剖结构指标与种源的冻害指数、冻害率的相关系数均没有达到显著水平。但是,通道种源翅荚木的CTR值显著大于其他种源,SR值显著小于其他种源,可以说明其抗寒性比较强。
|
|
植物对温度胁迫的抗性是自然选择的结果,在形态结构方面发生的适应性变化是一种相对稳定的遗传特性。这在苹果属(Malus)、柑桔属(Citrus)、白花泡桐(Paulownia fortunei)、白刺花(Sophora davidii)等植物上已经得到证实(黄以江等,1982;简令成,1986;苏金乐等,1993;钟广炎等,1994;李芳兰等,2006)。垂周壁形态、表皮毛长短和密度等常作为植物抗寒性的特征。陈清西等(1992)认为食用蕉(Musa)叶表皮所具的波浪状垂周壁,是其抗旱抗寒能力较强的表现。但翅荚木不同种源的垂周壁均呈U形、V形起伏,不同种源间没有明显差异。一般认为,表皮毛长而多是旱生植物叶的特点之一,这有助于反射阳光、防止水分蒸发,也有助于提高植物的抗寒能力。然而,虽然翅荚木不同种源的表皮毛长短和密度有所不同,但是与抗寒性并没有显著的相关性。
CTR和SR在多种林木上被用于指示抗寒性的强弱,一般CTR大小与抗寒性呈正相关,SR与抗寒性呈负相关。研究表明,通道种源翅荚木的CTR值显著大于其他种源,SR值显著小于其他种源,这与田间试验湖南通道种源抗寒性明显强于其他种源的结果相一致。说明翅荚木不同种源的CTR大小与抗寒性呈正相关,SR与抗寒性呈负相关。这与简令成(1986)、吴国良等(1998)、黄以江等(1982)和吴林等(2005)在其他林木上的研究结果也相一致。CTR值和SR值可作为鉴定翅荚木不同种源抗寒性的形态解剖指标。
电导率的测定作为室内评价植物抗寒性的指标在许多植物上得到运用(刘祖祺,1981;缴丽莉等,2006),一般认为植物组织电解质渗出率为50%时的温度正好与半致死温度一致(王荣富,1987)。植物组织的过冷温度的测定在国外已经陆续有相关的试验报道,国内由于试验条件的限制研究较少。余观夏等(2006)采用自制的试验装置测定了杨树无性系的超冷现象,证明杨树不同无性系的过冷温度与抗寒性有关。本研究根据植物组织过冷时有放热现象的特点,利用差示热量扫描仪测定了翅荚木不同种源叶片组织的过冷点温度。研究表明通道种源的叶片组织半致死温度和过冷点温度均低于其他种源,说明其抗寒性好于其他种源,这与田间试验结果相一致。
衡量林木对温度胁迫耐受性的指标有许多。一般生理生化指标受环境因素的影响较大,且操作上较为复杂。形态特征指标和生长发育特性的测定虽然简单,但一般不具备精确的可比性。植物抗寒基因研究在抗冻蛋白基因、减少膜伤害有关基因以及抗寒基因调节因子等方面也已取得一定进展。本文通过不同种源的形态解剖特性观察和生理生化分析,同时结合田间试验,可为翅荚木耐寒种源的筛选提供可靠的依据。
陈亮明, 陈永密, 张巧琴. 1997. 翅荚木引种栽培耐寒力的调查研究. 林业科技开发, (3): 37-38. |
陈清西, 廖镜思, 王明双, 等. 1992. 食用蕉若干品种类型叶片组织结构的比较观察. 福建农学院学报, 21(4): 406-412. |
陈永密, 张巧琴, 张玉石, 等. 1989. 珍稀树种——翅荚木. 福州: 福建教育出版社.
|
何义发, 严昌荣, 王柏泉. 1996. 珍稀树种——翅荚木在恩施山地引种试验初报. 湖北林业科技, (1): 24-26. |
侯伦灯, 李玉蕾, 李平宇, 等. 2001. 任豆树综合利用研究. 林业科学, 37(3): 139-143. DOI:10.3321/j.issn:1001-7488.2001.03.022 |
黄以江, 王宗清. 1982. 苹果属果树抗寒性的细胞学鉴定. 园艺学报, 9(3): 23-29. |
简令成. 1986. 不同种类桔柑叶片组织的细胞结构与抗寒性的关系. 园艺学报, 13(3): 163-168. |
缴丽莉, 路丙社, 白志英, 等. 2006. 四种园林树木抗寒性的比较分析. 园艺学报, 33(3): 667-670. DOI:10.3321/j.issn:0513-353X.2006.03.046 |
李芳兰, 包维楷, 刘俊华, 等. 2006. 岷江上游干旱河谷海拔梯度上白刺花叶片生态解剖特征研究. 应用生态学报, 17(2): 5-10. |
廖声熙, 张春华, 李立, 等. 2004. 印楝种源间抗寒能力比较研究. 林业科学研究, 17(2): 172-177. DOI:10.3321/j.issn:1001-1498.2004.02.006 |
刘世彪, 陈青, 胡正海. 2004. 7种番荔枝果树的叶片结构及其与抗寒性关系研究. 果树科学, 21(3): 241-246. DOI:10.3969/j.issn.1009-9980.2004.03.013 |
刘祖祺. 1981. 电导法鉴定柑桔耐寒性的试验. 南京农业大学学报, 2(6): 32. |
柳新红, 王军峰, 何小勇. 2005. 翅荚木种源引种苗期试验初报. 浙江林业科技, 25(5): 27-28. DOI:10.3969/j.issn.1001-3776.2005.05.008 |
荣文琛, 秦国峰, 吴天林, 等. 1999. 马尾松种源苗期抗寒性研究. 林业科学研究, 12(5): 534-538. DOI:10.3321/j.issn:1001-1498.1999.05.016 |
苏金乐, 程绍荣. 1993. 白花泡桐不同种源叶片比较解剖学研究. 河南农业大学学报, 27(1): 52-56. |
覃志刚, 邱进贤. 1991. 翅荚木引种栽培的研究. 四川林业科技, 13(1): 59-63. |
唐季林, 徐化成. 1989. 油松抗寒性与种源关系的研究. 北京林业大学学报, 11(1): 53-59. DOI:10.3321/j.issn:1000-1522.1989.01.001 |
王晶英, 敖红, 张杰, 等. 2003. 植物生理生化实验技术与原理. 哈尔滨: 东北林业大学出版社.
|
王利, 张东宁. 2002. 国内林木抗寒性研究的进展及展望. 山东林业科技, (1): 48-49. DOI:10.3969/j.issn.1002-2724.2002.01.024 |
王荣富. 1987. 植物抗寒指标的种类及其应用. 植物生理学通讯, (3): 49-55. |
吴国良, 常留印, 陈国秀. 1998. 核桃实生苗叶片性状与抗寒性关系. 植物学通报, 15(增刊): 111-113. |
吴林, 刘海广, 刘雅娟, 等. 2005. 越橘叶片组织结构及其与抗寒性的关系. 吉林农业大学学报, 27(1): 48-50, 54. DOI:10.3969/j.issn.1000-5684.2005.01.012 |
肖双燕, 李力. 1992. 翅荚木测树指标与冻害关系的初步研究. 江西林业科技, (6): 23-25. |
余观夏, 阮锡根, 封维忠, 等. 2006. 杨树无性系超冷现象分析. 南京林业大学学报:自然科学版, 30(2): 67-71. |
余光, 张亚宁. 1998. 我省引种的任豆木材纤维形态初步研究. 福建林业科技, 25(1): 31-33. DOI:10.3969/j.issn.1002-7351.1998.01.006 |
曾广腾, 龚期绳, 吴茂隆, 等. 2004. 翅荚木引种育苗试验及苗木生长规律研究. 江西林业科技, (2): 6-7. DOI:10.3969/j.issn.1006-2505.2004.02.002 |
张钢. 2005. 国外木本植物抗寒性测定方法综述. 世界林业研究, 18(5): 14-20. DOI:10.3969/j.issn.1001-4241.2005.05.004 |
张善江, Martin L Laps. 2005. 应用低温放热分析测定8个酒用葡萄新品种的抗寒性. 果树学报, 22(2): 121-124. |
钟广炎, 陈力耕. 1994. 柑桔种质资源抗寒性与叶片结构的关系. 中国柑桔, 23(2): 16-17. |
周志春, 李建民, 陈炳星, 等. 2003. 几种亚热带速生乡土阔叶树种的制浆特性评价. 中国造纸, 22(2): 8-12. DOI:10.3969/j.issn.0254-508X.2003.02.003 |
Ashworth E N, Lightner G W, Rowse D J. 1981. Evaluation of apricot flower bud hardiness using a computer-assisted method of thermal analysis. HortScience, 16(6): 754-756. |
Kang S K, Motosugi H, Yonemori K, et al. 1998. Supercooling characteristics of some deciduous fruit trees as related to water movement within the bud. Journal of Horticultural Science and Biotechnology, 73(2): 165-172. DOI:10.1080/14620316.1998.11510960 |
Warmund M R, George M F, Cumbie B G. 1990. Supercooling in 'Darrow' blackberry buds. Journal of the American Society for Horticulture Science, 113(3): 418-422. |