 2007, Vol. 43
2007, Vol. 43文章信息
- 李意德, 许涵, 陈德祥, 骆土寿, 莫锦华, 罗文, 陈焕强, 蒋忠亮.
- Li Yide, Xu Han, Chen Dexiang, Luo Tushou, Mo Jinhua, Luo Wen, Chen Huanqiang, Jiang Zhongliang.
- 从植物种群间联结性探讨生态种组与功能群划分——以尖峰岭热带低地雨林乔木层数据为例
- Discussing on the Ecological Species Groups and Functional Groups Division Basedon the Interspecific Association——A Case Study on the Arbor Layer Data in Tropical Lowland Rain Forest of Jianfenling, Hainan Island, China
- 林业科学, 2007, 43(4): 9-16.
- Scientia Silvae Sinicae, 2007, 43(4): 9-16.
-
文章历史
- 收稿日期:2006-03-17
-
作者相关文章


2. 海南尖峰岭国家级自然保护区 乐东 572542
2. Jianfengling National Nature Reserve, Hainan Province Ledong 572542
海南岛的热带低地雨林,也称热带常绿季雨林(胡玉佳等,1992;蒋有绪等,1991),其优势种青皮(Vatica mangachapoi)是东南亚地区森林表征科——龙脑香科(Dipterocarpaceae)青皮属(Vatica)植物,曾广布于海南岛各地(广东植物研究所,1975),现主要集中分布在尖峰岭、霸王岭、吊罗山、猕猴岭、万宁礼纪等地。作为国家二级重点保护植物的青皮,具有较广泛的经济利用价值,材质坚硬,不仅是建筑用材之一,也是重要的工业良材。由于具有树冠浓密、常绿、抗风和耐瘠薄的特点,近年也开始用于行道和景观绿化(周铁烽,2001)。青皮林群落学和林学特征等方面的研究已有一些报道(胡玉佳等,1988;梁淑群等,1994a;1994b;符国瑷等,1995;方洪等,2004;杨小波等,2005;李意德等,2006),也有学者对其植物区系(邢福武等,1993)和生物学特性(胡玉佳,1986)进行了研究,但对该森林植被类型的物种间关系以及更进一步的群落生态种组的划分尚未开展研究,在植物功能群的研究方面更是空白。本文试图通过对海南岛尖峰岭以青皮为主的热带低地雨林乔木层种群间联结的研究,探讨其生态种组和功能群的划分。
1 研究区概况尖峰岭林区位于海南岛西南部的乐东和东方两县交界处(18°23′—18°50′ N,108°36′—109°05′ E)。属低纬度热带岛屿季风气候,年均气温24.5 ℃,≥10 ℃年积温9 000 ℃,最冷月平均气温19.4 ℃,最热月平均气温27.3 ℃,干湿两季明显。成土母岩主要是含大量粉红色长石斑晶的似斑状花岗岩,土壤为砖黄壤。具有明显的气候、土壤和植被垂直带谱,地带性植被类型为热带常绿季雨林(黄全等,1986;蒋有绪等,1991;曾庆波等,1997;李意德等,2002;2006)。
研究地位于尖峰岭国家级自然保护区内的三分区,该区地处保护区西北部的核心区,海拔250~850 m。主要森林植被类型为以青皮为优势的湿润低地热带雨林,分布在海拔300~700 m的山体中部。由于海拔升高,降雨量比热带半落叶季雨林(海拔300 m以下)有较大幅度的增加,空气相对湿度也相应增大,虽有短暂的旱季,但干旱程度相对较轻。群落外貌为全年常绿,几乎没有落叶树种(黄全等,1986;胡玉佳等,1992)。
2 研究方法取样:采用典型取样法,从海拔约250 m处的热带半落叶季雨林至海拔约850 m的热带山地雨林,中间包括热带常绿季雨林,沿海拔梯度在林分结构完好的地段等间距设置23个样方,每个样方面积20 m×30 m,总调查面积为1.38 hm2。对样地内胸径≥12 cm的所有乔木树种进行调查,记录种名、胸径、树高等林分因子。23个样方的林分因子信息详见表 1(李意德等,2006)。
|
|

种间联结的测度:1)多物种间的总体关联性检验,采用Schluter (1984)提出的基于出现-不出现数据的方差比率来检验,其方差比率在独立性假设条件下期望值VR= 1,而VR>1时表示物种间表现出正的关联,VR<1时表示物种间存在负的净关联。2)利用2×2联列表(1个种对由“种A”和“种B”2个种组成,n值代表总样方数,a值代表 2个种均出现的样方数,b值代表 2个种均没出现的样方数,c值代表仅有种A出现的样方数,d值代表仅有种B出现的样方数),采用联结系数CA、种对共同出现百分率PC、点相关系数Φ、Ochiai指数QI以及χ2检验5种方法来检验成对物种间的联结性(Greig-Smith,1983;Pielou,1988;彭少麟,1996;王伯荪等,1996;王铮峰等,1997;黄世能等,2000;周先叶等,2000;骆土寿等,2005)。
1) χ2检验 采用Yates的连续校正公式计算
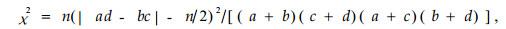
|
式中n为取样总数。当χ2<3.841时,种间联结独立;当χ2≥6.635时, 种间联结极显著; 当3.841≤χ2<6.635时,种间联结显著。χ2本身没有负值, 因此判定正、负联结的方法是:若a>(a+b)(a+c)/n(即ad>bc)时为正联结,反之为负联结。
2) 联结系数CA 用来进一步检验由χ2所测出的结果及说明种间联结程度。若ad≥bc, 计算公式为

|
若bc>ad且d≥a,则
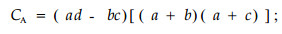
|
若bc>ad且d<a,则
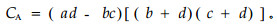
|
CA的值域为[-1,1]。其值越趋近于1,物种间正联结性越强;CA值越趋近于-1,物种间负联结性越强;CA值为0,物种间完全独立。
3) 共同出现百分率PC用来测度物种间正联结程度,其计算公式为
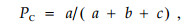
|
PC的值域为[0,1],其值越趋近于1,则表明该种对的正联结越紧密。
4) 点相关系数 Φ计算公式为
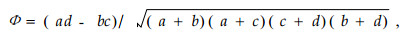
|
用于说明种间联结的程度,Φ为[-1,1]值域的有中心指数。
5) Ochiai指数计算公式为

|
该指数在“无关联”时等于0, “最大关联”时为1。
优势种群的聚类分析:为了进一步探索生态种组和功能群的划分,采用种群重要值对优势种进行聚类分析。种群重要值为样地中各种群的相对密度、相对优势度和相对频度之和(Cox, 1979),所有种群重要值之和为300,但本文采用相对重要值(即重要值除以3)。
生态种组和物种功能群的划分:依据优势物种之间的联结关系和聚类分析结果,初步划分不同的生态种组,并以此为依据进行功能群划分探讨。
数据处理:所有数据均在Excel 2003 (Microsoft Corporation)和Statistica 6.0 (StatSoft Inc.)等软件平台下进行分析和制图。
3 结果分析 3.1 种群重要值热带森林植物种类多,生物多样性高,为全面反映群落中优势种群和关键种群的地位和作用,根据样地调查数据,计算所调查样地的全部种群重要值,依据重要值大小排序,选取重要值在2.0(即相对重要值0.666 7)以上的32个种群做种间联结性分析(表 2)。
|
|
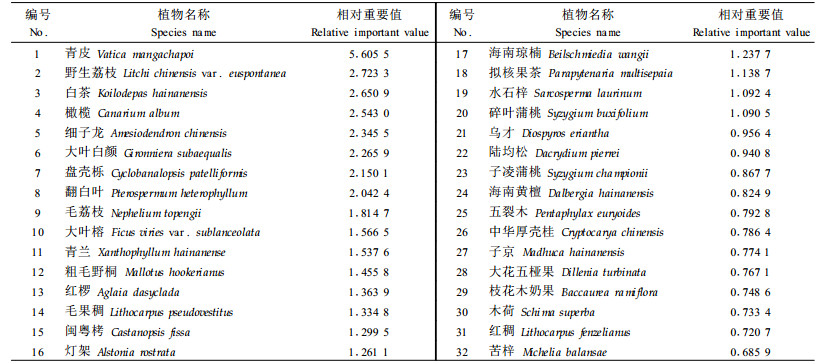
根据表 2所列举的32个重要值排前的种群,计算种群间的总体联结性,结果表明:VR=1.632 4>1,表明总体上种间存在一定的正联结, 其显著性统计量W为37.545 5,以自由度查相应χ2值, 即χ0.05(23)2= 35.172,χ0.95(23)2= 13.091,W值不落入χ0.05(23)2与χ0.95(23)2之间,且大于χ0.05(23)2值,表明32个种群间总体上正联结达显著水平,说明群落中主要种群间具有互利共存的关系。
3.2.2 主要种群间的联结从图 1可以看出:32个种群构成的496个种对中,48对具有强正联结性(0.6<CA≤1.0),占总种对数的9.8%,弱正联结(0.2<CA≤0.6)有101对,占20.36%,无联结(-0.2<CA≤0.2)有145对,占29.23%,弱负联结(-0.6<CA≤-0.2)有53对,占10.69%,强负联结(-1.0<CA≤-0.6)有149对,占30.04%,由此可见,具有正联结的种对占30.16%。

由于计算公式不同,共同出现百分率PC的计算结果与CA有很大的差异。由图 2可见:PC值在80%以上的为强正联结,在496个种对中,没有一对达到强正联结水平,PC值为60%~80%的次强正联结也仅有2个种对,它们是青皮与粗毛野桐、野生荔枝和海南琼楠。PC值为40%~60%的次弱正联结有30个种对;PC值20%~40%的弱正联结和20%以下的无联结的种对分别有137和327对。因此,热带林种群间联结性较弱,说明种群具有相对独立分布的特性(黄世能等,2000)。

|
图 2 优势种群间共同出现百分率PC半矩阵 Fig. 2 Half matrix of the percentage co-occurrence (PC) among the dominant populations |
点相关系数Φ的计算结果分级标准与CA相同。图 3结果表明:具有强正联结(Φ>0.6)的种对有7对,占总种对数的1.41%,弱正联结(Φ值0.2~0.6)的121对,占24.4%,无联结(Φ值-0.2~0.2)的占242对,占48.79%,弱负联结(Φ值-0.6~-0.2)的126对,占25.4%,强负联结(Φ值≤-0.6)的没有。具有强正联结的7个种对是:白茶与翻白叶、白茶与红椤、毛荔枝与大花五桠果、野生荔枝与海南琼楠、中华厚壳桂与子凌蒲桃、苦梓与五裂木、陆均松与闽粤栲。

|
图 3 优势种群间点相关系数Φ半矩阵 Fig. 3 Half matrix of the point correlation coefficients (Φ) among the dominant populations |
Ochai指数Qi的计算结果分级标准与PC相同。图 4结果表明:在496个种对中,没有一对达到强联结的水平,次强联结也仅有31个种对,占6.25%。次弱联结有106个种对,占21.37%;弱联结和无联结的种对分别有155和204对,占31.25%和41.13%。同样,Ochai指数Qi的计算结果也反映出了热带林种群间联结性较弱、种群具有相对独立分布的特性(黄世能等,2000)。

|
图 4 优势种群间Ochiai指数Qi半矩阵 Fig. 4 Half matrix of the Ochiai index (Qi) among the dominant populations |
根据χ2值查χ2表,当p>0.05时,即χ2<3.841,物种间无联结,为独立分布;当p<0.01时,χ2>6.635,物种间联结极显著;当0.01≤p≤0.05时,3.841≤χ2≤6.635,物种间联结显著。而种间的正负联结则由2×2联列表中物种A和物种B出现与否的具体情况而定(邓贤兰等;2003)。计算结果(图 5)表明:χ2>6.635,即物种间联结极显著的有2个种对,它们是野生荔枝与海南琼楠、毛荔枝与大花五桠果;而物种间联结显著的有大叶白颜与拟核果茶、青皮与粗毛野桐、白茶与野生荔枝、白茶与翻白叶、白茶与红椤、白茶与海南琼楠、拟核果茶与灯架、拟核果茶与五裂木、翻白叶与红椤、中华厚壳桂与苦梓、中华厚壳桂与子凌蒲桃、苦梓与五裂木、大花五桠果与大叶榕13个种对。大叶白颜与翻白叶、白茶与苦梓这2个种对虽然具有显著的联结关系,但为负联结;从这4个种的生物学特性来看,大叶白颜和苦梓是分布海拔较高的种,而翻白叶和白茶则分布海拔较低、且为耐热性较强的种类。

|
图 5 优势种群间联结的χ2检验半矩阵 Fig. 5 Half matrix of interspecific association χ2 test among the dominant populations |
利用这32个种群的相对密度、相对频度和相对优势度之和(即种群重要值)进行聚类分析,可以更好地理解这32个种群对环境的适应性和相似程度。
聚类结果清晰地表示出32个主要种群可分为4个类型(图 6),青皮作为热带低地雨林的主要种类,为独立种群,被单独列为一类,在尖峰岭西坡海拔300~750 m处(李意德等,2006)生长(但在尖峰岭的东坡最低海拔可达100 m),可在多种生境条件下生长,并且该种群更新情况良好(广东植物研究所,1975)。因此,青皮可认为是尖峰岭热带低地雨林的“生态关键种组”和“建群种组”。

|
图 6 青皮林主要种群聚类图 Fig. 6 Tree diagram of cluster analysis for 32 species 采用曼哈顿聚类系数和韦式法聚灶。 Using ward's method and city-block (manhattan) distances. |
组Ⅰ的种群有9个:野生荔枝、细子龙、红椤、水石梓、海南黄檀、枝花木奶果、海南琼楠、白茶和翻白叶,它们都是较为嗜热的种类,是热带低地雨林的次要种类(黄全等,1986;李意德等2006),但它们的分布并不像青皮那样广泛,只是在适宜的生境下才有较多的个体数量,同时在群落的层次结构中处于不同的地位,这些种群可称为低地雨林的“特征种群组”。
组Ⅱ的种群有9个:橄榄、毛荔枝、粗毛野桐、大花五桠果、乌才、大叶白颜、青兰、盘壳栎和大叶榕。这些种群一般在热带山地雨林中有一定地位(李意德,1997),但由于它们分布范围较广,且多为群落的中下层种类,因此它们在热带低地雨林中仍有主要的地位和作用,可称这类种群为低地雨林和山地雨林的“共有种群组”。
组Ⅲ的种群有12个:毛果稠、灯架、拟核果茶、闽粤栲、子凌蒲桃、木荷、五裂木、苦梓、红稠、碎叶蒲桃、陆均松和子京,它们是热带山地雨林典型的伴生种类(黄全等,1986;李意德,1997;曾庆波等,1997;李意德等,2002)。本次调查中,其出现在海拔较高的地段,如海拔650 m以上的样方中。相对于低地雨林而言,这些种群可称为“边缘种群组”。
对尖峰岭热带低地雨林的“生态关键种组”、“特征种群组”、“共有种群组”和“边缘种群组”的分类结果,与前述种群间的联结分析结果基本吻合。
3.4 物种功能群的划分功能群侧重于说明物种对生态过程和功能的作用,可在对生态种组分析的基础上进行辨识,将一些生态过程和功能相似的种类归为同一功能群。上文分析结果表明:青皮由于在群落中具有较高的重要值而成为尖峰岭地区热带低地雨林的优势种,或称为生态关键种;其他种群在生态种组的基础上进行划分,依据物种间的联结特性以及这些物种所处的海拔、微地形条件、在群落内的优势度和自身的生物学特性(如在林层垂直高度中所处的位置、嗜热程度等)等划分功能群,在此基础上尝试将32个主要物种划分为4个生态种组10个功能群(表 3),供后续研究参考。
|
|

各生态种组功能群辨识特征(或称为划分标准)如下:生态关键种组-建群物种在分布范围内占优势,种群结构正常,出现在林冠的各层次中;特征种组-亚优势物种在分布范围内占次优势,为上层乔木,但在不同的地形条件下其次优势地位有变化;特征种组-伴生物种的植株个体不一定是最大的,但一些种类的植株数量上有一定的优势,在林冠的上中下层均有,但以中层或下层为主;特征种组-嗜热物种在低地雨林的林窗或林缘出现较多;共有种组-霸王物种在群落中个体数量少,但个体特大,多突出林冠上层,对群落的结构与功能有较大的影响;共有种组-亚优势物种的海拔分布范围较广,个体较大,处于林冠的中上层;共有种组-伴生物种的海拔分布范围较广,个体较小,处于林冠的中下层,一些种的种群数量较多;边缘种组-伴生物种分布在海拔600 m以上处,个体较大,但种群较小,通常位于林冠的中上层;边缘种组-特殊物种一般多为分布在海拔700 m以上的山脊或山顶的特殊种类;边缘种组-更新物种一般出现在海拔500 m以上的林窗内。
4 结果与讨论 4.1 优势种群间联结对于群体而言,32个种群间总体上的正联结性反映了群落具有较强的稳定性,一般而言,稳定性是随着群落的演替进程而逐步加强的,它既反映在群落的结构稳定上,也反映在其物种组成的稳定性方面,群落越向顶极方向演替,其稳定性就越强(王伯荪等,1986;邓贤兰等,2003)。因此,尖峰岭自然保护区核心区内的青皮林原生性较强,结构和组成较为稳定,群落处于演替的后期阶段或动态平衡阶段。
虽然青皮林主要物种间总体上呈现出正联结的特性,但对于各物种之间而言,由于热带森林的物种多样性非常高(李意德,1995;李意德等,2002),组成群落的种类复杂,其物种间相遇的概率就很低,因此其种间联结性并不一定呈现出正联结的特性,而呈现出弱联结、零联结或负联结的可能性要大得多,这在一些热带林的种间联结性研究中已得到验证,表明热带森林的组成种群具有相对独立分布的特征(王峥峰等,1997;黄世能等,2000;戴小华等,2003;骆土寿等,2005),本研究结果也支持这一结论。正联结(或联结性强)的物种对所占的比例很少,反映了调查对象为具有成熟结构的森林生态系统,因为随着群落的正向演替,物种间的联结越具有无联结的趋势。
本研究具有正联结的种对中,建群种青皮只与海南黄檀、白茶、粗毛野桐、细子龙、盘壳栎少数几个种类具有一定的联结关系;而分布范围较广的大叶白颜与青兰、拟核果茶、苦梓、子凌蒲桃、灯架、水石梓、五裂木、陆均松、子京等多个种的联结性较强;白茶与野生荔枝、翻白叶、红椤、海南琼楠、大叶榕和海南黄檀等种群的关系密切;细子龙则与粗毛野桐的联结性较强。同时,还可以看出,一个物种对另一个物种有较高的联结值,不代表另一个物种对该物种也有较高的联结值,这与物种的生态位特征研究结论一致(李意德,1993)。
4.2 生态种组及功能群划分种间联结性指标可作为划分生态种组的重要依据之一,因为联结系数的高低反映了物种对环境资源的基本要求和生态系统组成结构特征。对32个主要种群进行聚类分析,可分成4大类(即4大生态种组):热带低地雨林的生态关键种青皮单独为一类,9个低地雨林的特征种类,9个热带低地雨林和山地雨林的共有种类,以及只在低地雨林与山地雨林的生态交错带出现的12个边缘种类。
根据物种的组成特征(物种间联结性、聚类分析)、生物学特性和分布特性等(广东植物研究所,1975;周铁烽,2001)划分了功能群。目前,对优势种群不明显且种类繁多的热带森林进行符合实际情况的功能群划分还有一定的难度,还需要引入其他数量指标(如森林小气候、物种的生理生态特征等)。另外,本文是根据热带低地雨林数据来划分功能群的,若以热带山地雨林数据为依据来划分,物种组成及其生物学特性又有不同,其结果应是另一种情况了。
戴小华, 余世孝, 练琚瑜. 2003. 海南岛霸王岭热带雨林的种间分离. 植物生态学报, 27(3): 380-387. DOI:10.3321/j.issn:1005-264X.2003.03.014 |
邓贤兰, 刘玉成, 吴杨. 2003. 井冈山自然保护区栲属群落优势种群的种间联结关系研究. 植物生态学报, 27(4): 531-536. DOI:10.3321/j.issn:1005-264X.2003.04.016 |
方洪, 李意德, 罗文, 等. 2004. 尖峰岭国家级自然保护区青皮林资源及其垂直分布特征. 热带林业, 32(4): 38, 43-46. |
符国瑷, 冯绍信. 1995. 海南五指山森林的垂直分布及其特征. 广西植物, 15(1): 57-69. |
广东植物研究所. 1975. 海南植物志:第4卷. 北京: 科学出版社.
|
胡玉佳, 李玉杏. 1992. 海南热带雨林. 广州: 广东高教出版社.
|
胡玉佳, 王寿松. 1988. 海南岛热带雨林优势种——青梅种群增长的矩阵模型. 生态学报, 8(2): 104-110. |
胡玉佳. 1986. 海南岛青梅种群生物学研究简报. 植物学通报, 4(1-2): 95-97. |
黄全, 李意德, 郑德璋, 等. 1986. 海南岛尖峰岭地区热带植被生态系列的研究. 植物生态学与地植物学学报, 10(2): 90-105. |
黄世能, 李意德, 骆土寿, 等. 2000. 海南岛尖峰岭次生热带山地雨林树种间的联结动态. 植物生态学报, 24(5): 569-574. DOI:10.3321/j.issn:1005-264X.2000.05.010 |
蒋有绪, 卢俊培. 1991. 中国海南岛尖峰岭热带林生态系统. 北京: 科学出版社.
|
李意德, 陈步峰, 周光益, 等. 2002. 中国海南岛热带森林及其生物多样性保护研究. 北京: 中国林业出版社.
|
李意德, 方洪, 罗文, 等. 2006. 海南尖峰岭国家级保护区青皮林资源与乔木层群落学特征. 林业科学, 42(1): 1-6. |
李意德. 1993. 海南岛尖峰岭热带山地雨林主要种群生态位特征研究. 林业科学研究, 7(1): 78-85. |
李意德. 1995. 海南岛热带森林的变迁及生物多样性的保护对策. 林业科学研究, 8(4): 455-461. DOI:10.3321/j.issn:1001-1498.1995.04.003 |
李意德. 1997. 海南岛尖峰岭热带山地雨林的群落结构特征. 热带亚热带植物学报, 5(1): 18-26. |
梁淑群, 林英, 杨小波, 等. 1994a. 海南万宁礼纪青梅林(续). 海南大学学报:自然科学版, 12(1): 14-19. |
梁淑群, 林英, 杨小波, 等. 1994b. 海南万宁礼纪青梅林(续完). 海南大学学报:自然科学版, 12(2): 129-135. |
骆土寿, 李意德, 陈德祥, 等. 2005. 海南岛鸡毛松人工林群落种间联结性研究. 生态学杂志, 24(6): 591-594. DOI:10.3321/j.issn:1000-4890.2005.06.001 |
彭少麟. 1996. 南亚热带森林动态. 北京: 科学出版社.
|
王伯荪, 余世孝, 彭少麟, 等. 1996. 植物群落学实验手册. 广州: 广东高等教育出版社.
|
王铮峰, 安树青, Campell D G, 等. 1997. 热带山地雨林种间联结的测定. 内蒙古大学学报:自然科学版, 23(3): 400-406. |
邢福武, 李泽贤, 吴德邻. 1993. 海南岛南部甘什岭植物区系的初步研究. 植物研究, 13(3): 227-242. |
杨小波, 吴庆书, 李跃烈, 等. 2005. 海南北部地区热带雨林的组成特征. 林业科学, 41(3): 19-24. DOI:10.3321/j.issn:1001-7488.2005.03.004 |
曾庆波, 李意德, 陈步峰, 等. 1997. 热带森林生态系统研究与管理. 北京: 中国林业出版社.
|
周铁烽. 2001. 中国热带主要经济树木栽培技术. 北京: 中国林业出版社.
|
周先叶, 王伯荪, 李鸣光, 等. 2000. 广东黑石顶自然保护区森林次生演替过程中群落的种间联结性分析. 植物生态学报, 24(3): 332-339. DOI:10.3321/j.issn:1005-264X.2000.03.015 |
Cox G W. 1979.普通生态学实验手册.蒋有绪, 译.北京: 科学出版社
|
Greig-Smith P. 1983. Quantitative Plant Ecology. Oxford: Blackwell, 54-104.
|
Pielou E C. 1988.数学生态学.卢泽愚, 译.北京: 科学出版社
|
Schluter D A. 1984. Variance test for detecting species associations, with some example applications. Ecology, 65(3): 998-1005. DOI:10.2307/1938071 |