 2007, Vol. 43
2007, Vol. 43文章信息
- 杨玉盛, 陈光水, 王义祥, 谢锦升, 杨少红, 钟羡芳.
- Yang Yusheng, Chen Guangshui, Wang Yixiang, Xie Jinsheng, Yang Shaohong, Zhong Xianfang.
- 格氏栲人工林和杉木人工林碳吸存与碳平衡
- Carbon Sequestration and Balance in Castanopsis kawakamii and Cunninghamia lanceolata Plantations in Subtropical China
- 林业科学, 2007, 43(3): 113-117.
- Scientia Silvae Sinicae, 2007, 43(3): 113-117.
-
文章历史
- 收稿日期:2006-01-16
-
作者相关文章


2. 福建省农业科学研究院 福州 350002;
3. 福建农林大学林学院 福州 350003
2. Fujian Academe of Agricultural Sciences Fuzhou 350002;
3. School of Forestry, Fujian Agricuture and Forestry University Fuzhou 350002
森林是陆地生态系统中最大的碳库,其碳贮量占全球陆地总碳贮量的46%(Watson et al., 2000)。因而,开展森林生态系统碳平衡动态研究,对于科学预测森林对全球碳平衡和气候变化的作用具有重要意义,并已成为国际陆地碳循环研究的热点。目前国外有关国家、区域及生态系统水平的森林碳平衡的研究较多,大多集中于温带森林和热带森林(Grace et al., 1995; Goulden et al., 1996; Yamamoto et al., 2001)。近10年来我国在森林碳平衡方面亦已有一定研究。在国家尺度上,方精云等(2001)对我国50年来森林碳库、平均碳密度和CO2源汇功能变化进行了研究;刘国华等(2000)、周玉荣等(2000)、王效科等(2001)也分别利用森林资源清查资料从不同角度对我国森林生态系统的碳贮量和碳密度进行了分析。在区域尺度上,张娜等(2003)、焦秀梅等(2005)、曹军等(2002)和张德全等(2002)分别对长白山自然保护区、福建省、湖南省、海南省和山东省森林碳储量动态进行了研究;在不同森林生态系统水平上,李意德等(1998)对海南岛尖峰岭热带山地雨林生态系统的碳储量、CO2的排放以及生态系统碳平衡进行了研究;方晰等(2002)对会同森林定位站第二代杉木(Cunninghamia lanceolata)人工林速生阶段的碳密度、碳储量及其空间分布特征和碳平衡进行了研究;刘建军等(2003)对秦岭定位站油松(Pinus tabulaeformis)、锐齿栎(Quercus aliena)等主要森林类型的土壤呼吸及土壤碳循环进行了研究;王金叶等(2000)对青海云杉(Picea crassifolia)林的碳汇功能及其碳平衡进行了研究;方精云等(2006)对北京东灵山3种温带森林生态系统大储量、碳通量和碳平衡进行了研究。
我国南方集体林区是我国商品林或用材林的重要基地,如何在提供木材的同时最大限度地发挥其碳吸存潜力已成为我国南方林区林业可持续发展的必然趋势,亦是缓解国际社会对我国CO2限制压力的重要举措之一。目前国际上已提出碳人工林(carbon plantation)的概念。虽然碳人工林目前尚无确切定义,但可理解为是一种超越以木材收获为主的传统经营目标,以同时追求木材收获和碳吸存效益为双重目标的新型人工林经营模式(Onigkeit et al., 2000)。如何在提供木材的同时最大限度地发挥其碳吸存潜力已成为我国南方林区林业可持续发展的必然趋势,亦是缓解国际社会对我国CO2限制压力的重要举措之一。碳平衡是评价不同树种碳汇大小的重要途径。本文在定位研究基础上,对格氏栲(Castanopsis kawakamii)和杉木人工林碳吸存与碳平衡进行分析比较,为我国南方林区碳汇树种选择提供借鉴。
1 试验地概况试验地位于福建省三明市莘口镇。格氏栲人工林(CK)和杉木人工林(CF)的郁闭度分别为0.95和0.85,平均树高分别为18.9和21.9 m,平均胸径分别为24.2和23.3 cm,密度分别为875和1 117株·hm-2,蓄积量分别为431 m3·hm-2(杨玉盛等,2003a; 2003b;2004a; 2004b)。格氏栲人工林土壤密度为1.10 g·cm-3, 有机质、全N、全P、水解N、速效P含量分别为29.8 g·kg-1、1.12 g·kg-1、0.31 g·kg-1、115.2 mg·kg-1和5.92 mg·kg-1。杉木人工林土壤密度为1.20 g·cm-3,有机质、全N、全P、水解N和速效P含量分别为29.5 g·kg-1、1.12 g·kg-1、0.29 g·kg-1、110.3 mg·kg-1和4.69 mg·kg-1(杨玉盛等,2003a; 2003b;2004a; 2004b)。格氏栲人工林和杉木人工林均为1967年在格氏栲天然林采伐迹地上营造,调查时(2002年)年龄均为36年生。
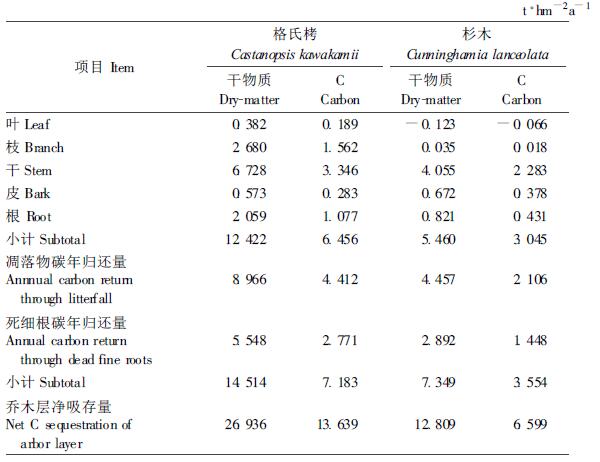
2 研究方法 2.1 植株碳测定和乔木层碳年净吸存量计算乔木层各器官、凋落物各组分和枯死细根碳含量采用元素分析仪(ELEMEMTAR Vario EL Ⅲ)测定。乔木层生物量碳净增量为各器官生物量净增量与各器官相应碳浓度的乘积之和。采用相对生长法推算36年生时格氏栲和杉木人工林乔木层各器官的生物量年净增量(杨玉盛等,2003c),即建立各器官生物量与胸径、树高的相关方程,通过测定当年胸径、树高和下一年胸径、树高,分别代入生物量方程,求得各器官生物量之差即为各器官生物量年净增量。乔木层枯落物(包括凋落物和枯死细根)碳年归还量为枯落物各组分归还量与相应组分碳浓度的乘积之和。凋落物和枯死细根年归还量的测定方法参见文献(杨玉盛等,2003a; 2003b)。生态系统各组分的碳含量数据为叶494 g·kg-1、枝583 g·kg-1、干497 g·kg-1、皮495 g·kg-1、根508 g·kg-1(杨玉盛等,2006a)。本文中凋落物和枯死细根年归还量均为2002年数据,见表 1。乔木层碳年净吸存量为乔木层生物量碳净增量和枯落物碳归还量之和。
|
|
土壤异养呼吸(为无根土壤呼吸和枯枝落叶层呼吸之和)年通量为2002年定位观测数据,无根土壤呼吸和枯枝落叶层呼吸具体测定和数据见文献(杨玉盛等,2006b)。
2.3 生态系统碳平衡计算(方晰等,2002;方精云等,2006)生态系统碳收入项为I=NPP=BI+L,其中NPP为生态系统中乔木层年净初级生产量中的碳量,BI为乔木层年生物量增量中的碳量,L为乔木层通过枯落物(包括凋落物和枯死细根)归还的碳量(本计算中忽略了林下植被归还量)。
生态系统碳支出项为O=Rh,其中Rh为生态系统异养呼吸,在不考虑动物的取食作用时,Rh即相当于土壤(包括矿质土壤和枯枝落叶层)的异养呼吸。
则生态系统碳平衡公式为ΔC=NEP=I-O=NPP-Rh=BI+L-Rh,其中ΔC为生态系统碳年增量,NEP为净生态系统生产量中的碳量,即为生态系统年净碳汇。NEP或净碳汇是生态系统和大气之间CO2交换的直接度量, 也是评价生态系统尺度碳源/汇的直接指标(方精云等,2006)。
方晰等(2002)和方精云等(2006)曾提出类似的碳平衡计算公式,但方晰等的碳平衡支出项直接以土壤呼吸代替土壤异养呼吸,方精云等的碳平衡收入项中则考虑了生物量增量和地上凋落物,而忽略了地下凋落物部分。
3 结果 3.1 乔木层碳年净吸存量36年生格氏栲人工林乔木层碳年净吸存量是36年生杉木林的2.1倍;2种人工林乔木层净碳吸存量中生物量净增量和枯落物(凋落物与死细根)归还量各约占50%(表 1),表明在此生长阶段枯落物归还量和乔木层生物量净增量对乔木层碳汇贡献相当。格氏栲人工林干材(干+皮)生物量增量占乔木层净碳吸存量的26.6%,远低于杉木人工林(40.3%);格氏栲人工林的枝和根生物量增量占乔木层净碳吸存量的比例(11.5%和7.9%)则均高于杉木人工林(0.3%和6.5%)。这与2个树种生物学特性差异有关。格氏栲和杉木人工林的地上和地下枯落物碳年归还量均约占年碳总归还量的60%和40%左右,表明中亚热带枯落物归还中,虽然细根归还占有重要位置,但凋落物仍然占据主要地位。
3.2 生态系统碳平衡36年生格氏栲人工林和杉木人工林净碳汇(净生态系统生产量NEP)分别为7.656和3.615 t C·hm-2 a-1,即每年可分别净固定CO2 28.072和13.255 t·hm-2a-1,均表现出“碳汇功能”。而格氏栲人工林的净碳汇是杉木人工林的2.1倍(表 2)。
|
|

格氏栲人工林乔木层碳年净吸存量(13.639 t·hm-2a-1)与我国热带林(14.55 t·hm-2a-1)和常绿阔叶林(12.29 t·hm-2a-1)的平均水平接近(周玉荣等,2000),但高于尖峰岭热带山地雨林(8.615 t·hm-2 a-1)(李意德等,1998)和亚热带35年生青冈(Cyclobalanopsis glauca)常绿阔叶林(7.754 t·hm-2a-1)(李铭红等,1996)。杉木人工林乔木层碳年净吸存量(6.599 t·hm-2a-1)则小于我国暖性针叶林的平均水平(7.97 t·hm-2a-1)(周玉荣等,2000)及本试验地33年生福建柏(Fokienia hodginsii)人工林(9.907 t·hm-2a-1)(何宗明等,2003),但高于会同10年生杉木人工林(5.496 t·hm-2a-1)(方晰等,2002),而与本试验地27年生杉木林(6.691 t·hm-2a-1)相近(杨玉盛等,2003c)。本研究中格氏栲乔木层碳年吸存量高于杉木人工林的,这与格氏栲人工林乔木层的碳积累速率大且高碳积累持续时间长(杉木人工林乔木层碳吸存的数量成熟龄为20年,而格氏栲人工林的则至33年生时仍未达数量成熟)有关(杨玉盛等,2006a)。
目前我国有关森林土壤异养呼吸的研究很少。目前仅有李意德等(1998)和方精云等(2006)进行过相关报道。周玉荣等(2000)虽然报道了我国森林碳平衡状况,但其中土壤呼吸和土壤异养呼吸数据借用国际研究数据。本研究通过挖壕沟法排除根系呼吸而得到土壤异养呼吸(杨玉盛等,2006b),格氏栲人工林土壤异养呼吸年通量与热带林(6.93 t C·hm-2a-1)(周玉荣等,2000)和尖峰岭热带山地雨林(7.352 t C·hm-2a-1)(李意德等,1998)相近,但高于常绿落叶阔叶林(4.90 t C·hm-2a-1)、硬叶常绿阔叶林(3.92 t C·hm-2 a-1)(周玉荣等,2000)、白桦(Betula platyphylla)林(2.78 t C·hm-2a-1)及辽东栎(Quercus liaotungensis)林(3.49 t C·hm-2 a-1)(方精云等,2006);杉木人工林则低于暖性针叶林(3.75 t C·hm-2a-1)(周玉荣等,2000)和油松林(1.81 t C·hm-2a-1)(方精云等,2006)。
格氏栲人工林净碳汇与热带林(7.68 t·hm-2a-1)和常绿阔叶林(7.28 t·hm-2a-1)的平均水平接近(周玉荣等,2000),但高于澳大利亚北部热带萨瓦纳森林(3.8 t·hm-2a-1)(Chen et al., 2003)、欧洲温带落叶林(2~5.9 t·hm-2a-1)(Goulden et al., 1996; Malhi et al., 1999)、亚马逊热带森林(1.00 t·hm-2a-1)(Grace et al., 1995)、中国尖峰岭热带山地雨林原始林(0.373 t·hm-2a-1)(李意德等,1998)、马萨诸塞州温带落叶混交林(1.4~2.8 t·hm-2a-1)(Guris et al., 2002)及日本温带落叶林(1.28 t·hm-2a-1)(Yamamoto et al., 2001)。杉木人工林净碳汇小于俄罗斯西伯利亚东部200年生欧洲赤松(Pinus sylvestris)林(4.4 t·hm-2a-1)(Wirth et al., 2002),但大于中国兴安落叶松(Larix gmelinii)林(2.65 t·hm-2 a-1)(蒋延玲等,2001)。本研究中人工林净碳汇与不同地区森林的差异可能与气候条件、林龄和树种生物学特性差异有关。方晰等(2002)曾报道会同10年生杉木林表现为净碳源(-0.427 t·hm-2a-1),但其在碳平衡计算中直接利用土壤呼吸代替异养呼吸。方精云等(2006)曾报道白桦林、辽东栎林和油松林的净碳汇(NEP)分别为0.95、-0.29和4.08 t·hm-2a-1,但其在碳平衡计算时NPP组成中忽略了地下净生产力部分。
格氏栲和杉木人工林碳平衡2个分量间的比率(土壤异养呼吸/乔木层净碳吸存量,即碳释放/碳固定)相近(分别为0.44和0.45),表明格氏栲和杉木人工林间净碳汇的差异主要由碳通量的(碳循环强度)差异引起,即格氏栲人工林的碳收入项和碳支出项均成比例地大于杉木人工林,从而使格氏栲人工林的净碳汇大于杉木人工林。
目前,有关碳人工林树种选择所涉及的因素较为复杂,受到树种的生物学特性、市场条件(木材、碳信用价格)、成本和利率条件等诸多因素的影响(Onigkeit et al., 2000)。碳人工林的经营目标是达到木材收益和碳吸存收益两者综合效益的最大化,因而如何在两者效益间达到最佳的平衡是目前研究的难点。单从碳吸存数量的角度考虑,作为一种碳人工林的适宜树种,首先是其生长速率快,即能在较短时间内达到其最大碳积累量。其次是该树种碳人工林具有较高水平的最大碳积累量。其三是该碳人工林轮伐期越长(轮伐期取决于木材和碳吸存综合效益达到最大化的时间),则其碳吸存效益越高。其四是该碳人工林在采伐后进入社会经济系统的碳吸存量要高(即木材收获中的碳积累量)。满足以上4个方面才能保证人工林拥有较高的碳吸存效益。
本研究中,36年生格氏栲人工林的碳贮量远高于36年生的杉木人工林,表明在36年中格氏栲人工林的生长速率快于杉木人工林;且36年生时格氏栲人工林的净碳汇仍远大于杉木人工林,可以预测在今后短期内格氏栲人工林的碳积累量仍将高于杉木人工林的。假定格氏栲和杉木人工林均在36年生时采伐(轮伐期相同),则格氏栲人工林收获时木材中的碳积累量(110.4 t·hm-2)亦高于杉木人工林(83.4 t·hm-2)(杨玉盛,2006a)。因而,格氏栲人工林无论在森林立地内,抑或在社会经济系统内,其碳吸存效益均高于杉木人工林。单从碳吸存效益的角度考虑,格氏栲是一种比杉木更优良的碳人工林树种。
曹军, 张镱锂, 刘燕华. 2002. 近20年海南岛森林生态系统碳储量变化. 地理研究, 21(5): 551-560. DOI:10.3321/j.issn:1000-0585.2002.05.003 |
方精云, 陈安平. 2001. 中国森林植被碳库动态变化及其意义. 植物学报, 43(9): 967-973. DOI:10.3321/j.issn:1672-9072.2001.09.014 |
方精云, 刘国华, 朱彪, 等. 2006. 北京东灵山三种温带森林生态系统的碳循环. 中国科学D辑:地球科学, 36(6): 533-543. |
方晰, 田大伦, 项文化, 等. 2002. 第二代杉木中幼林生态系统碳动态与平衡. 中南林学院学报, 22(1): 2-6. |
何宗明, 李丽红, 王义祥, 等. 2003. 33年生福建柏人工林碳库与碳吸存研究. 山地学报, 21(3): 298-303. DOI:10.3969/j.issn.1008-2786.2003.03.007 |
蒋延玲, 周广胜. 2001. 兴安落叶松林碳平衡和全球变化影响研究. 应用生态学报, 12(4): 481-484. DOI:10.3321/j.issn:1001-9332.2001.04.001 |
焦秀梅, 项文化, 田大伦. 2005. 湖南省森林植被的碳贮量及其地理分布规律. 中南林学院学报, 25(1): 4-8. DOI:10.3969/j.issn.1673-923X.2005.01.001 |
李铭红, 于明坚, 陈启 NFDA1, 等. 1996. 青冈常绿阔叶林的碳素动态. 生态学报, 16(6): 645-651. |
李意德, 吴仲民, 曾庆波, 等. 1998. 尖峰岭热带山地雨林生态系统碳平衡的初步研究. 生态学报, 18(4): 371-378. |
刘国华, 傅伯杰, 方精云. 2000. 中国森林碳动态及其对全球碳平衡的贡献. 生态学报, 20(5): 733-740. DOI:10.3321/j.issn:1000-0933.2000.05.004 |
刘建军, 王得祥, 雷瑞德, 等. 2003. 秦岭天然油松、锐齿栎林地土壤呼吸与CO2释放. 林业科学, 39(2): 8-13. |
王金叶, 车克钧, 蒋志荣. 2000. 祁连山青海云杉林碳平衡研究. 西北林学院学报, 15(1): 9-14. DOI:10.3969/j.issn.1001-7461.2000.01.002 |
王效科, 冯宗炜, 欧阳志云. 2001. 中国森林生态系统的植物碳储量和碳密度研究. 应用生态学报, 12(1): 13-16. DOI:10.3321/j.issn:1001-9332.2001.01.003 |
杨玉盛, 陈光水, 林鹏, 等. 2003a. 格氏栲天然林与人工林细根生物量、季节动态及净生产力. 生态学报, 23(9): 1719-1730. |
杨玉盛, 陈光水, 王义祥, 等. 2006a. 格氏栲人工林和杉木人工林碳库及分配. 林业科学, 42(10): 43-47. |
杨玉盛, 陈光水, 谢锦升, 等. 2006b. 格氏栲天然林与人工林土壤异养呼吸特性及动态. 土壤学报, 43(1): 53-61. |
杨玉盛, 郭剑芬, 林鹏, 等. 2004a. 格氏栲天然林与人工林枯枝落叶层碳库及养分库. 生态学报, 24(2): 359-367. |
杨玉盛, 林鹏, 郭剑芬, 等. 2003b. 格氏栲天然林与人工林凋落物数量、养分归还及凋落叶分解. 生态学报, 23(7): 1278-1289. |
杨玉盛, 刘艳丽, 陈光水, 等. 2004b. 格氏栲天然林与人工林土壤非保护性有机C含量及分配. 生态学报, 24(1): 1-8. |
杨玉盛, 谢锦升, 王义祥, 等. 2003c. 杉木观光木混交林C库与C吸存. 北京林业大学学报, 25(5): 10-14. |
张德全, 桑卫国, 李日峰, 等. 2002. 山东省森林有机碳储量及其动态的研究. 植物生态学报, 26(增刊): 93-97. |
张娜, 于贵瑞, 赵士洞, 等. 2003. 长白山自然保护区生态系统碳平衡研究. 环境科学, 24(1): 24-32. |
周玉荣, 于振良, 赵士洞. 2000. 我国主要森林生态系统碳库和碳平衡. 植物生态学报, 24(5): 518-522. DOI:10.3321/j.issn:1005-264X.2000.05.002 |
Chen X Y, Hutley L B, Eamus D. 2003. Carbon balance of a tropical savanna of northern Australia. Oecologia, 137: 405-416. DOI:10.1007/s00442-003-1358-5 |
Curtis P S, Hanson P J, Bolstad P, et al. 2002. Biometric and eddy-covariance based estimates of annual carbon storage in five eastern North American deciduous forests. Agricultural and Forest Meteorology, 113: 3-19. DOI:10.1016/S0168-1923(02)00099-0 |
Goulden M L, Munger J W, Fan S M, et al. 1996. Measurements of carbon acquisition by long-term eddy covariance:methods and critical evaluation of accuracy. Global Change Biology, 2: 169-182. DOI:10.1111/j.1365-2486.1996.tb00070.x |
Grace J, Lloyd J, Miranda A C, et al. 1995. Carbon dioxide uptake by an undisturbed tropical rain forest in South-West Amazonia 1992-1993. Science, 270: 778-780. DOI:10.1126/science.270.5237.778 |
Malhi Y, Baldocchi D D, Jarvis P G. 1999. The carbon balance of tropical, temperate and boreal forests. Plant, Cell & Environment, 22(6): 715-740. |
Onigkeit J, Sonntag M, Alcamo J. 2000. Carbon Plantations in the IMAGE Model-Model Description and Scenarios. WZ Ⅲ Report No. P0003. Center for Environmental Systems Research, University of Kassel, Germany
|
Watson R T, NobleI R, Bolin B, et al.2000. IPCC Special Report on Land Use.Land Use Change and Forestry
|
Yamamoto S, Saigusa N, Murayama S, et al.2001. Long-term results of flux measurement from a temperate deciduous forest site(Takeyama).Proceedings of International Work-shop for Advanced Flux Network and Flux Evaluation.Sapporo: ASAHI Printing CO.Ltd, 5-10
|