 2007, Vol. 43
2007, Vol. 43文章信息
- 黄金燕, 周世强, 谭迎春, 周小平, 王鹏彦, 张和民.
- Huang Jinyan, Zhou Shiqiang, Tan Yingchun, Zhou Xiaoping, Wang Pengyan, Zhang Hemin.
- 卧龙自然保护区大熊猫栖息地植物群落多样性研究:丰富度、物种多样性指数和均匀度
- Study on the Species Diversity of Plant Community in the Giant Panda Habitat of Wolong Natural Reserve:Species Richness, Species Diversity and Evenness
- 林业科学, 2007, 43(3): 73-78.
- Scientia Silvae Sinicae, 2007, 43(3): 73-78.
-
文章历史
- 收稿日期:2005-04-25
-
作者相关文章


野生动物保护管理的本质在于其栖息地的保护,包括原生生境的巡护和监测、退化生境的恢复与重建、基因交流和领域扩展的走廊建设等。评估野生动物栖息地的质量与数量特征,笔者认为除进行定性的概念性描述外,生境结构的定量数据是评价栖息地质量性状的重要指标,也是恢复退化生境的参考依据、建立自然保护区的理论基础、评估保护管理成效的有效参数。从生态学角度分析,野生动物栖息地的质量结构主要由食物、水和隐蔽物三大因子组成(Bailey,1984)。食物因子因野生动物的食性差异而不同,对食肉动物来说,与其伴生的野生动物是其食物资源;对素食动物而言,栖息地中的植物组分是其食物来源。水的理化特性对野生动物的生长发育和繁殖更新具有重要的作用,因而形成了不同野生动物在不同的生长发育期、不同的生长季节中饮用不同水质的觅食对策。隐蔽物除岩洞、土穴等外,野生动物主要利用各种生态系统,包括森林、草原、湿地等自然植被类型。因此,分析植物群落的多样性是研究和评估野生动物栖息地的重要手段。
群落多样性是指生物群落在组成、结构、功能和动态方面表现出的丰富多彩的差异,在一定的景观或区域内,景观的异质性是人们首先感知的,而景观的异质性格局是由群落的多样性所决定的。因此,群落多样性研究是认识景观的组织结构水平和功能状态的基础(马克平,1994;马克平等,1995)。在分析卧龙自然保护区大熊猫栖息地的植物种类组成、区系地理成分和基本植物群落类型的基础上(周世强等,2003a;2003b),本文拟从植物群落的丰富度、均匀度和物种多样性指数的现状及其分布格局与生境的关系,以及多样性与植物生长型的关系等方面进行分析,以期为大熊猫栖息地植物群落性质的认识、保护管理及评价提供参考。
1 自然概况与研究方法 1.1 自然概况卧龙自然保护区位于四川省汶川县西南部,邛崃山系的东南坡,岷江上游,102°52′—103°24′E、30°45′—31°25′N,东西横贯60 km,南北跨越63 km,总面积2 000 km2,是我国最大的自然保护区之一,主要保护大熊猫(Ailuropoda melanoleuca)、金丝猴(Pygathrix roxellanae)、牛羚(Budorcas taxicolor)、珙桐(Davidia involucrata)、光叶珙桐(Davidia involucrata var. vilmoriniana)、水青树(Tetracentron sinense)、四川红杉(Larix mastersiana)等珍稀濒危动植物以及整个高山生态系统。
卧龙地处四川盆地向青藏高原过渡的高山峡谷地带,地势由西北向东南急剧递减。由于新构造运动的抬升作用及河流的侵蚀影响,区内山高谷深,相对高差悬殊。西北的四姑娘山海拔高达6 250 m,东部木江坪海拔仅1 150 m,两地相距48 km,相对高差达5 100 m。区内主要河流有皮条河、正河、西河和中河,河流两侧发育许多各级支流,形成树枝状水系,河谷呈“V”形状,落差较大,具有丰富的水电资源。
该区属青藏高原气候区,夏季凉爽多雨,冬季寒冷干燥。平均气温9.8 ℃,最低气温(1月)-1.7 ℃,最高气温(7月)17 ℃,年日照时数926.7 h,年降雨量1 800 mm,蒸发量873.9 mm,相对湿度80%以上。
根据张万儒(1983)对卧龙自然保护区森林土壤及其垂直分布规律的研究,本区发育有从亚热带到冰缘环境的各种土壤类型。从河谷到山顶,主要土壤类型垂直分布为:山地黄壤、山地黄棕壤、山地暗棕壤、山地棕色暗针叶林土、亚高山草甸土、高山草甸土和高山寒漠土。
该区主要分布有常绿阔叶林、常绿落叶阔叶混交林、落叶阔叶林、温性针叶林、温性针阔混交林、寒温性针叶林、耐寒灌丛、高山草甸和高山流石滩稀疏植被等9个植被类型(卧龙自然保护区管理局等,1987)。
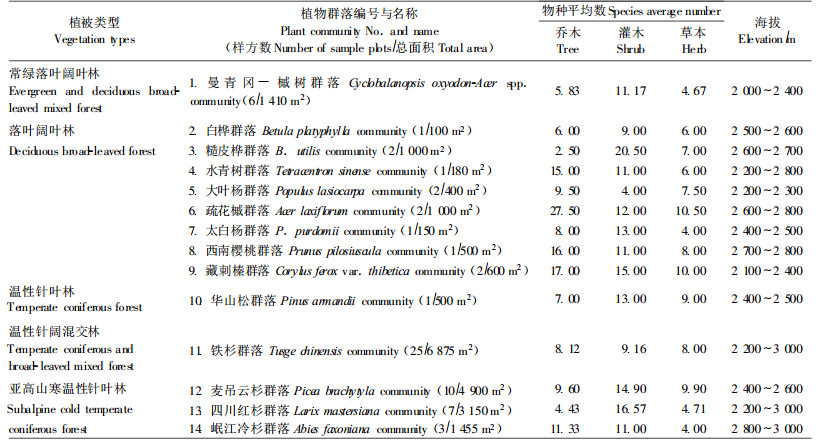
1.2 研究方法 1.2.1 样地概况根据卧龙野生大熊猫种群数量的地理分布和栖息地利用情况,在海拔2 000~3 000 m大熊猫分布范围内设置了64个植被调查样方(面积100~500 m2)进行群落多样性分析。大熊猫栖息地的主要植物群落类型及其物种数量与分布范围等列于表 1。
|
|
参见文献(周世强等,2003a;2003b)。
1.2.3 植物群落多样性的测度群落的物种多样性是反应其组织水平的重要生态学特征之一,它是群落结构类型、组织水平、发展阶段、稳定程度和生境差异的体现(黄忠良等,2000)。群落的多样性指数由丰富度(绝对密度)和均一性(相对密度)组成(岳天祥,2001),是其种数、个体总数和均匀程度的综合概念(Whittaker,1969),是把物种数和均匀度结合起来的一个单一的统计量(马克平,1994)。为了综合评价卧龙自然保护区野生大熊猫栖息地的质量结构,采用了物种丰富度指数、物种多样性指数和均匀度指数测度群落的多样性(马克平等,1995;史作民等,2002)。各指标的计算方法及公式分述于下:
Shannon-Wiener指数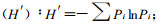
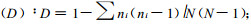
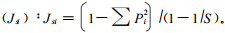
地形是表征调查样地内热量与水分含量多寡的定性地理参数,它包括坡向、坡度和坡位3个指标,而坡位常用海拔值进行说明。
2.1.1 坡向对大熊猫栖息地植物群落物种多样性的影响在一定的海拔和坡度范围内,坡向影响着水热的重新分配和组合,成为影响植物分布与生长的主要地形因素。数据分析表明(图 1),坡向对区内植物群落物种多样性分布格局有一定的影响,在物种丰富度、物种多样性指数和均匀度指数上的反应是不同的:不同坡向的物种丰富度指数S变化最显著,其趋势为西>北>东>南,Shannon-Wiener指数H′和Simpson指数D均呈现出西>南>北>东的递降格局,而均匀度指数Jsi则是南>西>东>北的动态变化规律。

|
图 1 不同坡向的大熊猫栖息地多样性指数 Fig. 1 The diversity indices of Giant Panda habitat in different aspects of slope  下同。The same below. 下同。The same below.
|
依坡度的物种多样性指数变化规律来看(图 2),物种多样性最高的坡度范围为21~30°,但方差分析(S:F=0.318<F0.05=2.53;H′:F=0.586<F0.05=2.53;D:F=0.537<F0.05=2.53;Jsi:F=0.586<F0.05=2.53)表明0~5°、6~20°、21~3 0°、31~40°和≥41°各坡度等级之间的多样性指数差异不显著,这说明坡度并不是影响卧龙野生大熊猫栖息地生物多样性的关键因子。

|
图 2 不同坡度的大熊猫栖息地多样性指数 Fig. 2 The diversity indices of Giant Panda habitat in the different slopes |
坡位生态因子的作用常用海拔变化的影响来说明。在特定的区域范围内,海拔是影响植物群落物种分布与组成变化的决定性因素。海拔的变化导致生境条件的不同,从而在海拔梯度上形成生物多样性差异,进而影响到植物群落的空间分布格局(马克平等,1995)。由图 3可知植物群落的物种多样性在海拔梯度上的变化:随着海拔的升高,物种丰富度指数快速上升,至海拔2 101~2 200 m的曼青冈-槭树群落和藏刺榛群落时其上升速度减缓,在海拔2 301~ 2 400 m的曼青冈-槭树群落、水青树群落、藏刺榛群落、铁杉群落和四川红杉群落处达到最大值后下降,在海拔2 601~3 000 m范围因分布着针叶林群落、铁杉群落、四川红杉群落和岷江冷杉群落和处于演替进程中的落叶阔叶树种群落如糙皮桦群落、水青树群落、疏花槭群落、西南樱桃群落,其物种丰富度出现波动;Shannon-Wiener指数H′随海拔升高逐渐增加,但在海拔2 101 m~2 400 m的曼青冈-槭树群落、大叶杨群落、水青树群落、藏刺榛群落、铁杉群落和四川红杉群落达到最大值后开始下降至平稳;而均匀度指数Jsi和Simpson指数随海拔的上升其变化较一致,最初微升,然后减少,但最后于2 401~ 2 500 m海拔后趋于平稳,即针阔混交林和针叶林群落物种中个体数目或蓄积量趋于微小变化,同时这一区域因原始针叶林与处于演替进程中的落叶阔叶树种群落的镶嵌分布,故其物种多样性的垂直分布格局出现微弱变化。比较而言,物种丰富度随海拔的变化与贡嘎山东坡海拔1 600~2 600 m范围的变化趋势类似(沈泽昊等,2001),而海拔2 100 m以上均匀度指数Jsi和Simpson指数随海拔的变化趋势与白龙江上游地区森林植物群落类似(郭正刚等,2003)。

|
图 3 不同海拔大熊猫栖息地植物群落多样性指数 Fig. 3 The diversity indices of plant communities in Giant Panda habitat at different elevations |
植物群落类型不同,其结构、功能和景观都存在差异,这些差异主要是因为生境的异质性导致组成植物群落的具有不同生态生物学特性的物种构成及其个体数量的不同,从而形成不同的群落物种多样性空间分布结构,且不同的植物群落构成了特定的植被景观类型。
卧龙自然保护区大熊猫栖息地的不同植被类型的多样性指数值除物种丰富度S变化为温性针叶林>落叶阔叶林>常绿落叶阔叶林>亚高山针叶林>温性针阔混交林外,H′、D、Jsi基本表现出相似的动态格局:常绿落叶阔叶林>温性针阔混交林>落叶阔叶林>温性针叶林>亚高山寒温性针叶林(表 2)。这是特定的地理环境、坡度、坡向和海拔等条件综合影响所致。
|
|

区内14个不同植物群落类型的多样性测度指标除物种丰富度外均呈现出基本一致的变化趋势(图 4)。在不同的植物群落中,如4号水青树群落其分布区在地形地势上为台地,坡度缓,土壤肥厚,水热条件好,则其群落的多样性指数较高;而5号大叶杨群落由于林冠郁闭度大、林层结构简单、林下植物组成种类较少等原因,其物种多样性较低。这些指数的变化都较好地反映了不同植物群落类型在物种组成方面的差异。

|
图 4 大熊猫栖息地不同植物群落类型的多样性指数 Fig. 4 The diversity indices of the different plant communities types in the Giant Panda habitat |
植物物种生长型是表征植物群落外貌特征和垂直结构的重要指标(马克平等,1995),生长型及其物种组成不同决定了植物群落的空间结构特征。按照Whittaker等(1975)的分类系统,这里以植物群落在空间结构上的最主要层次乔木层、灌木层和草本层等3个类型为研究对象分析卧龙自然保护区大熊猫栖息地物种多样性与植物生长型的关系。一般来说,植物群落的垂直结构主要受地带性气候所确立的水热组合的影响,但在一个特定的区域地带性气候区内,植物群落垂直层次结构则受群落所处的海拔、坡度、坡向、坡位等物理微环境和群落的物种组成、发育阶段、生活史对策等的影响,相反,植物群落特定的垂直层次结构及其物种多样性的动态直接反映了该群落内垂直生态梯度。
在研究区域内,这14个植物群落由于受微环境和建群种构成及发育特性的影响,其植物群落生长型的生物多样性变化极具特点(见表 1和图 5~7)。

|
图 5 大熊猫栖息地不同植物群落乔木层的多样性指数 Fig. 5 The diversity indices of tree layer of different plant communities in Giant Panda habitat |

|
图 6 大熊猫栖息地不同植物群落灌木层的多样性指数 Fig. 6 The diversity indices of shrub layer of different plant communities in Giant Panda habitat |

|
图 7 大熊猫栖息地不同植物群落草本层的多样性指数 Fig. 7 The diversity indices of herb layer of different plant communities in Giant Panda habitat |
乔、灌、草各层的物种丰富度存在一定的差异,在总体水平上表现为灌木层最大,乔木层次之,草本层最小,但42.9%群落中乔木层的物种丰富度大于灌木层,64.3%群落中乔木层的物种丰富度大于草本层,92.9%群落(5号群落因乔木层郁闭度过大除外)灌木层中的物种丰富度大于草本层;在不同群落中乔木层物种丰富度指数的变异幅度最大,灌木层次之,草本层最小。故乔木层或灌木层的物种丰富度在大熊猫栖息地不同植物群落中占优势,但因群落类型不同其各层片的物种组成相异。在不同生长型中,物种多样性指数和均匀度指数的变化趋势均大体相同,且其变化幅度在不同群落上有一定差异;均匀度指数变化幅度为灌木层大于乔木层,草本层最小,Simpson指数变化程度与均匀度指数近似,Shannon-Wiener指数的变化程度相对小一些,草本层大于灌木层和乔木层,而乔木层和灌木层的变化幅度近似。在同一生长型中,物种多样性指数和均匀度指数变化大体一致,其中Simpson指数和均匀度指数变化规律除草本层的1号群落外其它群落更接近一致,而Shannon-Wiener指数变化规律主要在草本层与Simpson指数变化略有不同,同时,物种多样性指数与均匀度指数的变化规律和物种丰富度指数变化在乔木层中基本一致,在灌木层中多数一致,而在草本层中部分一致。
以上大熊猫栖息地群落生物多样性分布格局不仅与各地形因子的水热条件组合分异有关,而且受制于组成群落的物种的生态生物学特性和地形地势的影响。由于卧龙地区具有沟壑纵横、河谷深切的峡谷地貌特征,由此导致微地形的水热分布差异明显,进而形成与地形因子变异一致的区域性生物多样性分布格局和珍稀濒危物种的“自然庇护所”(卧龙自然保护区管理局等,1987)。
3 结论卧龙自然保护区大熊猫栖息地的不同植被类型的物种多样性指数值H′、D和均匀度指数Jsi变化趋势基本一致:常绿落叶阔叶林>温性针阔混交林>落叶阔叶林>温性针叶林>亚高山针叶林,而物种丰富度S变化为温性针叶林>落叶阔叶林>常绿落叶阔叶林>亚高山针叶林>温性针阔混交林。在不同的植物群落中,由于地形地势、坡度、土壤、水热条件等原因,其物种多样性不同,但其多样性测度指标变化均呈现出基本一致的趋势。
在不同的海拔,其发育着不同特征的植物群落。随海拔的升高,大熊猫栖息地物种丰富度、多样性指数和均匀度变化先增加,然后下降,最后趋于平稳。
坡度不是影响卧龙野生大熊猫栖息地生物多样性的关键因子。坡向对群落多样性有影响。不同的坡向其物种丰富度指数S变化为西>北>东>南,Shannon-Wiener指数H′和Sim pson指数D呈现西>南>北>东的递降格局,均匀度指数Jsi呈现南>西>东>北的递降格局。
在同一或不同的生长型中,物种多样性指数和均匀度指数变化大体一致。42.9%群落中乔木层的物种数多于灌木层,64.3%群落中乔木层的物种数多于草本层,92.9%群落灌木层中的物种数多于草本层;落叶针叶林乔木层树种丰富度小于灌木层和草本层,但其灌木层物种丰富度大于草本层。
郭正刚, 刘慧霞, 孙学刚, 等. 2003. 白龙江上游地区森林植物群落物种多样性的研究. 植物生态学报, 27(3): 388-395. DOI:10.3321/j.issn:1005-264X.2003.03.015 |
黄忠良, 孔国辉, 何道泉. 2000. 鼎湖山植物群落多样性的研究. 生态学报, 20(2): 193-198. |
马克平. 1994. 生物群落多样性的测度方法//钱迎倩, 马克平, 生物多样性研究的原理与方法. 北京: 中国科学技术出版社, 141-165
|
马克平, 黄建辉, 于顺利, 等. 1995. 北京东灵山地区植物群落多样性的研究. 生态学报, 15(3): 268-277. DOI:10.3321/j.issn:1000-0933.1995.03.006 |
沈泽昊, 方精云, 刘增力, 等. 2001. 贡嘎山东坡植被垂直带谱的物种多样性格局分析. 植物生态学报, 25(6): 721-732. DOI:10.3321/j.issn:1005-264X.2001.06.013 |
史作民, 程瑞梅, 刘世荣, 等. 2002. 宝天曼植物群落物种多样性研究. 林业科学, 38(6): 17-23. DOI:10.3321/j.issn:1001-7488.2002.06.004 |
卧龙自然保护区管理局, 南充师范学院, 四川省林业厅保护处. 1987. 卧龙植被及资源植物. 成都: 四川科学技术出版社, 11-194.
|
岳天祥. 2001. 生物多样性研究及其问题. 生态学报, 21(3): 462-467. DOI:10.3321/j.issn:1000-0933.2001.03.019 |
张万儒. 1983. 卧龙自然保护区的森林土壤及其垂直分布规律. 林业科学, 19(3): 254-268. |
周世强, 黄金燕, 谭迎春, 等. 2003a. 卧龙大熊猫栖息地植物群落多样性研究Ⅰ.植物群落的基本特征. 四川林业科技, 24(2): 6-11. |
周世强, 黄金燕, 谭迎春, 等. 2003b. 卧龙自然保护区大熊猫栖息地植物群落多样性研究Ⅱ.植物群落的聚类分析. 四川林勘设计, (3): 16-20. |
Bailey J A. 1984. Principles of wildlife management. John Wiley & Sons, 196-252.
|
Whittaker R H. 1969. Evolution of diversity in plant communities. Ecology, 50: 417-428. |
Whittaker R H, Niering W A. 1975. Vegetation of the Santa Catalina Mountain, Arizona. V. Biomass, production, and diversity along the elevation gradient. Ecology, 56: 771-790. DOI:10.2307/1936291 |