 2007, Vol. 43
2007, Vol. 43文章信息
- 李新岗, 刘惠霞, 侯慧波, 高文海.
- Li Xingang, Liu Huixia, Hou Huibo, Gao Wenhai.
- 油松球果对外源茉莉酸甲酯和虫害诱导的生化反应
- Response of Chinese Pine Cones to Induction of Exogenous Methyl Jasmonate and Gravitarmata margarotana Larvae
- 林业科学, 2007, 43(3): 66-72.
- Scientia Silvae Sinicae, 2007, 43(3): 66-72.
-
文章历史
- 收稿日期:2005-10-08
-
作者相关文章


2. 西北农林科技大学植物保护学院 杨凌 712100
2. College of Plant Protection, Northwest A & F University Yangling 712100
害虫取食会诱导植物产生一系列的生理生化反应,并激活植物防御基因,引起植物代谢过程重新配置,使植物产生有毒化合物、抗营养和抗消化酶类、妨碍行为化合物及驱避剂等直接防御;并诱导植物挥发物吸引寄生性、捕食性天敌进行间接防御(桂连友等,2004; Kessler et al., 2001;2002)。这些反应对植物是有益的,是目前植物诱导抗性和植物-昆虫-天敌相互关系的研究热点(Dicke et al., 2000;Dicke,2000;Zhu-Salzman et al., 2005)。但是诱导植物产生负防御,也称诱导敏感性(induced susceptibility)(Dicke,2000),即虫害引起另一种害虫聚集和危害,这种诱导对植物是不利的。同时,对被引诱者来讲,一面临着竞争者和天敌的存在(刘芳等,2003);二面临食物质量下降、抗性物质增加(娄永根等,1997;Kessler et al., 2002)。
昆虫取食后,通过系统素或细胞膜信号级联反应,经过茉莉酸途径激活植物防御基因。所以,外源茉莉酸(jasmonic acid,JA)、茉莉酸甲酯(methyl jasmonate,MJA)等也可诱导与虫害类似的植物反应(Kessler et al., 2001;2002;Thaler,1999)。萜类是针叶树组成防御和诱导防御的特异性化学物质,挥发性萜类则作为昆虫寄主选择的信号物质。在针叶树中,虫害和MJA处理都可诱导化学防御物质的从头合成(Heijari et al., 2005;Martin et al., 2002;2003)。
油松球果小卷蛾(Gravitarmata margarotana)和松果梢斑螟(Dioryctria pryeri)在我国北方严重危害油松(Pinus tabulaeformis)球果,特别是以生产种子为目的的良种基地。油松球果小卷蛾先期危害,引起松果梢斑螟的聚集和严重危害, 表现为梢斑螟幼虫趋向小卷蛾危害的球果取食(李新岗等,2006),而成虫则趋向受害的球果产卵(李宽胜,1992)。为了探讨小卷蛾危害引起油松球果的负防御以及梢斑螟幼虫的适应机制,本文研究了外源MJ A和小卷蛾危害引起油松球果挥发物、抗营养(多酚氧化酶PPO)和抗消化酶类(蛋白酶抑制剂PI)以及球果游离氨基酸、松脂等成分的变化。
1 材料与方法 1.1 供试材料油松样树选择球果害虫危害较重的陕西陇县八渡油松种子园和没有球果害虫危害的西北农林科技大学林学院校园。陇县油松种子园建于1985年,海拔约1 500 m,树高3~5 m,375株·hm-2,郁闭度0.4~0.5,坡度20~30°;林学院校园海拔500 m,油松树高3~5 m,密度5 m×6 m。样品采集时间为小卷蛾幼虫危害初期,即梢斑螟越冬幼虫转移期的5月上中旬(2005年)。
在种子园随机选择3株油松作为样树,确定树冠中部半阳面为采样部位,采用顶空吸附法吸附样品。球果分析样品也是分别采自2处样地。吸附样品和球果样品用冰壶带回实验室,置于-20 ℃冰箱中待用。
接虫用的2~3龄小卷蛾幼虫,于2005年5月上旬采自陇县油松种子园的虫害球果,带回实验室待用。
1.2 试剂与仪器二氯甲烷(色谱纯,美国TEDIA公司);吸附剂XAD2(Ambeliate公司);茉莉酸甲酯(M J A)(Aldrich公司);α-胰凝乳蛋白酶,P-甲苯磺酰-L-精氨酸甲酯(TAME),异抗坏血酸盐,N-琥珀酰-L-苯丙氨酸-ρ-硝基苯和三乙胺(Sigma公司)。
TraceDSQ气相色谱-质谱仪(GC-MS)(美国Thermo-Finnigan产品),SIGMA 3K30型高速冷冻离心机,Beckman DU@70型荧光分光光度计和121MB型氨基酸分析仪,脂肪测定系统(瑞典Soxtec System HT 1043 Extraction Unit)。
1.3 球果挥发成分分析 1.3.1 球果挥发物的采集选择油松健康球果、小卷蛾危害球果或MJA处理球果;吸附剂XAD2活化后烘干装柱,每柱(Φ6 mm×12 mm)装XAD2150 mg;球果样品带1 0~15 cm长的新梢,用Tedler采样袋在样树上活体吸附5 h(每吸附50 min间隔10 min),空气流速1 L·min-1,吸附样品置于-20 ℃环境中保存;分析前用3 mL二氯甲烷洗脱(Dormont et al., 1998),洗脱液用于GC-MS分析。
1.3.2 GC-MS分析条件利用GC-MS分析进行挥发物的鉴定,色谱柱利用SE-54毛细管柱(30 m×0.25 mmID,膜厚0.25 μm),载气为氦气;流速1 mL·min-1,进样1 μL,分流比50:1。起始温度40 ℃,保持2.5 min,以8 ℃·min-1升到250 ℃。质谱条件:EI离子源,电离能70 eV。各成分通过与谱库(NIST2002版)标准化合物的质谱图核对分析并兼顾色谱保持时间后定性,根据峰面积归一化法进行定量。
1.4 球果多酚氧化酶(PPO)和蛋白酶抑制剂(PI)活性测定 1.4.1 样品处理于2005年5月分别在陇县油松种子园和校园进行采样。每株样树选择3个侧枝,1个侧枝为虫害球果和健康球果采样,2个侧枝为MJA处理枝。2年生球果(油松种子园)采用MJA暴露法处理,选用40 cm×50 cm的聚乙烯塑料袋将新稍和球果套上,每袋放1棕色小瓶,内装MJA 200 μL和95%乙醇2 mL浸入脱脂棉,对照只装2 mL 95%乙醇脱脂棉。当年生球果(校园)采用MJA喷雾法处理,每个结果枝喷100 mL MJA水溶液(含MJA 200 μL和95%乙醇2 mL),对照为喷蒸馏水100 mL(含2 mL 95%乙醇)和不处理2种(桂连友等,2005)。部分MJA处理球果也用于顶空吸附试验。
油松球果接虫试验在没有虫害干扰的校园油松上进行。选择半阳面的2年生球果若干,每果接虫1头(2~3龄小卷蛾幼虫),2 d后接虫成功的球果作为接虫受害球果采样。
1.4.2 样品分析球果PPO和PI活性采用分光光度法测定(桂连友等,200 5)。1年生球果先将其等分成2~3块,每个样品包括2~3个球果的材料;2年生球果在球果中部的鳞片或球果中部的内部组织采样。
1) PPO活性测定 酶液制备:取待测样品0.25 g,加不溶性聚乙烯吡咯酮(pvp)0.3 g、石英砂2 g及适量预冷的pH 5.6柠檬酸缓冲液,在冰浴中研磨,并用上述缓冲液定溶到10 mL,再置于4 ℃的冰箱中浸提12 h(搅拌几次);接着4 000 r·min-1离心15 min,上清液即为酶液。活性测定:采用分光光度计测定。取酶液1 mL,加反应混合液3 mL(混合液配置为0.1 mol·L-1柠檬酸缓冲液(pH 5.6):0.1%脯氨酸:1%临苯二酚10:2 :3)。然后在37 ℃恒温水浴中保温10 min,立即加1 mol·L-1偏磷酸3 mL终止反应。于460 nm波长处,用10 mm比色皿比色。空白管反应混合液中的临苯二酚用缓冲液代替。酶活性以每克样品每分钟E460增加0.1为1活性单位。
2) PI活性测定 酶液制备:取样品0.25 g,加0.25 g pvp,在液氮中研碎;再加入1.2 mL缓冲液(100 mmol·L-1 Tris-HCl,1%异抗坏血酸盐,pH 7.8)混匀后,4 000 r·min-1×10 min离心2次,得上清液。活性测定:选用α-胰凝乳蛋白酶(牛乳)为蛋白酶,采用荧光分光光度计测定PI活性。加入200 μL研磨液、750 μL缓冲液(100 mL三乙胺,200 mmol·L-1 CaCl2,pH 7.8)和25 μL蛋白酶液(α-胰凝乳蛋白酶200 μg·mL-1,2.5 mmol·L-1 HCl)混匀,放置20 min(约25 ℃),再加入500 μL基质溶液(12 mmol·L-1 N-琥珀酰-L-苯丙氨酸-ρ-硝基苯胺、20%乙醇、160 mmol·L-1三乙胺,pH 7.8),在25 ℃下放置30 min,于405 nm波长下进行吸光测定(反应物ρ-硝基丙氨酸的最大吸收波长),测定其吸光度。空白为不加研磨的上清液。以固定时间30 min的反应混合物测定OD值,酶活性以每克每分钟E405减少0.01为1活性单位。计算公式为:

|
并以对照组PI活性标记为0(桂连友等,2005)。
1.5 球果游离氨基酸和其他成分分析在陇县油松种子园,于5月中旬选择2年生健康球果和小卷蛾危害球果,切碎烘干,用于粗脂肪的提取和含水量的测试;鲜球果切碎后进行脱脂处理(罗玉萍等,2001),脱脂样品用蒸馏水定容后,在氨基酸分析仪上进行游离氨基酸分析。粗脂肪采用脂肪测定系统测定,球果含水量采用烘干称重法确定。
1.6 数据处理统计分析软件为ExcelStat (Installshield Software Corporation)。采用单因素(ANOVA)F检验(Fisher LSD post-hoc tests)比较不同处理间的差异显著性。
2 结果与分析 2.1 诱导的球果挥发成分变化 2.1.1 外源MJA和虫害诱导当年生球果枝中挥发成分比较表 1比较了外源MJA喷雾法处理2 d后,2个校园油松株系当年生球果枝挥发成分的变化。MJA处理后,单萜挥发成分总量明显增加,根据GC峰面积比较,处理样品单萜挥发成分平均增加1.58倍。从各成分含量变化看,2个株系处理后α-蒎烯和莰烯含量下降,而β-水芹烯、β-蒎烯和D-柠檬烯则增加,但株系2处理后β-月桂烯含量明显增加,株系1却没有变化。2个株系挥发物各组分的含量和处理后各组分的增减百分数有较大的差异,这种差异主要是株系及其生理状态造成的,也可能与试验的环境有关。
|
|
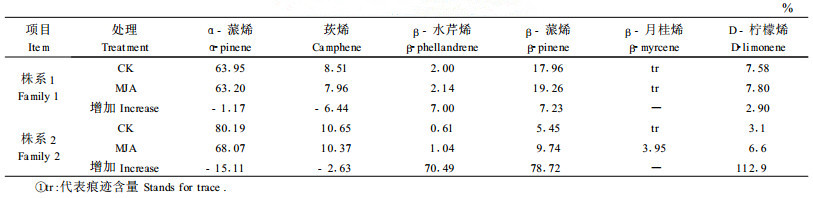
表 2比较了林间小卷蛾虫害球果枝和健康球果枝挥发成分的变化。结果显示:虫害球果枝和健康球果枝相比,α-蒎烯和莰烯含量下降,而β-蒎烯、β-月桂烯和D-柠檬烯含量增加,β-水芹烯则表现为很低水平没有变化;比较结果发现,虫害球果枝单萜总量增加2.49倍(据G C峰面积),高于MJA处理与对照的增加量,这可能与虫害诱导强度大、时间长有关。与未受球果害虫危害的油松样品(表 1)相比,除β-月桂烯含量明显增加、β-水芹烯处于很低水平外,各成分的变化趋势一致,但表 1和表 2中健康球果(对照)各挥发成分含量有很大的差异,这除了由于环境和发育进度差异外,主要与林间油松长期处于球果害虫的危害,球果已被虫害系统诱导有关。
|
|

在陇县油松种子园,外源MJA处理2 d后,2年生球果枝的α-蒎烯、莰烯、β-水芹烯、β-蒎烯和D-柠檬烯含量下降,而β-月桂烯含量则增加(表 3);单萜挥发成分总量增加了2.63倍(GC峰面积比较)。以2年生健康球果和虫害球果单萜挥发成分总量比较,虫害球果枝单萜挥发成分总量增加了5倍,且α-蒎烯、莰烯、β-水芹烯和β-蒎烯含量降低;而β-月桂烯和D-柠檬烯含量则增加(表 3)。除D-柠檬烯含量变化不同外,其他成分的变化与外源MJA诱导的变化一致。这可能是诱导时间和强度不同造成的,因为虫害诱导时间一般在7 d左右,而外源诱导仅2 d。2种诱导引起单萜挥发成分增加的倍数比较,虫害球果也明显大。
|
|

同样是油松种子园样树,2年生健康球果枝挥发成分总量是1年生健康球果枝挥发成分总量的5.12倍(GC峰面积比较),而2年生虫害球果枝挥发成分总量是1年生虫害球果枝挥发成分总量的4.54~10倍。所以推断,2年生虫害球果对天敌的作用显著高于1年生虫害球果,林间天敌的寄生情况调查也支持了这种推断(李宽胜,1992)。
2.2 外源MJA和虫害诱导球果多酚氧化酶(PPO)和蛋白酶抑制剂(PI)的变化 2.2.1 外源MJA和虫害诱导当年生球果中PPO和PI的变化在没有油松球果害虫危害情况下(校园),油松当年生球果经MJA喷雾处理2 d后,球果多酚氧化酶和蛋白酶抑制剂活性(用α-胰凝乳蛋白酶活力下降表示)显著增加,而喷蒸馏水100 mL(含2 mL 95%乙醇)和不处理的对照间无显著性差异(表 4)。
|
|

在林间条件下,健康球果和虫害球果的PPO和α-胰凝乳蛋白酶活性没有显著性差异。这可能是由于相邻枝当年生球果或同一枝的2年生球果受害后,通过系统诱导导致健康球果的PPO和PI活性也系统增加。
2.2.2 人工接虫和虫害诱导2年生球果中PPO和PI的变化在没有油松球果害虫危害情况下(校园),油松2年生球果经人工接虫处理2 d后,虫害球果中部虫果面PPO活性显著高于健康球果和虫害球果的健果面(表 5),诱导的系统性并不明显;陇县种子园(林间)采集的2年生健康球果和虫害球果则表现为明显的诱导系统性。PI活性在2年生球果上,表现出明显的诱导系统性,2种处理的虫害球果α-胰凝乳蛋白酶活性显著低于相应健康球果(表 5),即虫害球果PI活性显著高于健康球果。这可能是种子园球果害虫连年危害,长时间的系统诱导,因而造成林间样品PI活性要比无害虫危害的校园样品高得多。相比之下,就PPO和PI来说,当年生球果的诱导反应明显,而2年生球果的诱导反应不敏感,从而出现不同组织诱导反应的差异(Van Dam et al., 2001)。
|
|

表 6结果表明,在梢斑螟幼虫转移期,球果受害后,17种游离氨基酸含量及其总含量都发生了变化。其中游离氨基酸总含量下降1.04%,苏氨酸和颉氨酸含量没有变化;脯氨酸、精氨酸、组氨酸和蛋氨酸含量下降,降低幅度为11.0%~39.1%;而含量增加的氨基酸有11种,增长幅度在3.36%~80.5%之间,其中谷氨酸、酪氨酸和苯丙氨酸增长的比例最多。
|
|
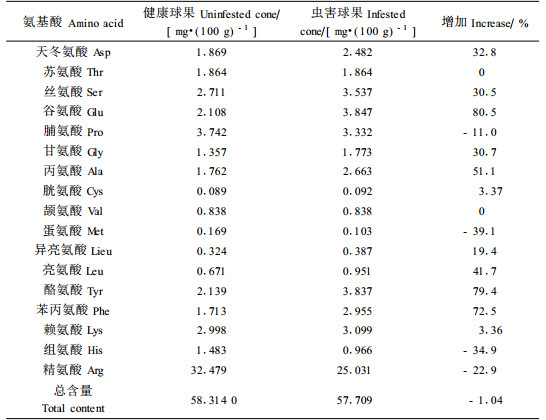
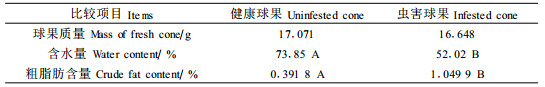
表 7比较了松果梢斑螟幼虫转移期健康球果和虫害球果的松脂(用粗脂肪表示)和水分含量,结果显示:虫害球果的含水量显著降低,但粗脂肪的含量显著增加。含水量的降低、松脂含量的增加,表明球果的营养质量在下降(康乐,1995;李镇宇等,1998)。
|
|
油松1年生和2年生球果枝主要挥发物均为6种单萜成分,即α-蒎烯、莰烯、β-水芹烯、β-蒎烯、β-月桂烯和D-柠檬烯。小卷蛾虫害诱导后,1年生和2年生球果枝单萜挥发成分种类没有变化,只是含量发生了变化,并且挥发成分总量明显增加;1年生球果受害后,α-蒎烯和莰烯含量减少,β-蒎烯、β-月桂烯和D-柠檬烯含量增加,但β-水芹烯则一直保持微量水平;2年生球果受害后,α-蒎烯、莰烯、β-水芹烯和β-蒎烯含量下降,而β-月桂烯和D-柠檬烯含量增加;外源MJA诱导后,与受害球果挥发成分有类似的变化。球果受害或MJA处理后,2年生球果与1年生球果挥发物总量有很大的差异,结合林间天敌调查数据(李宽胜,1992)推测,2年生虫害球果挥发物对天敌有更强的引诱作用,但具体机制及哪种挥发成分起作用还需进一步研究确定。
油松球果6种挥发成分,与火炬松(Pinus taeda)2年生球果(5种)(Hanula et al., 1985),以及火炬松庖锈病(Cronartium quercuum)的锈庖(7种)(Fatzinger et al., 1985),瑞士五针松(P. cembra)、华山松(P. armandii)等球果(7~8种)相近(Dormont et al., 1997;1998),主要成分都包括α-蒎烯、β-水芹烯、β-蒎烯、β-月桂烯和D-柠檬烯,因树种和采样时间不同,各成分及其异构体的含量变化较大。针叶树球果受害及外源激素诱导后,PPO和PI变化的研究还没有报道,但针叶树其他组织中PPO(姚延梼等,1998)以及虫害诱导针叶化学成分的变化已有报道,本研究结果与油松针叶受害后的成分变化一致(李镇宇等,1998)。
外源MJA处理和小卷蛾幼虫危害后,均可引起1年生和2年生球果PPO和PI活性增加,并且1年生球果对2种诱导的反应更敏感,系统性更强;而2年生球果PPO的系统诱导则较弱,PI的显著增加可能是2年期间球果虫害诱导的累积结果。虫害后引起17种游离氨基酸含量发生变化,但各氨基酸对球果营养质量的作用还不清楚。尽管如此,2年生球果受害后,PPO和PI活性增加、含水量下降、松脂含量增加,这都说明球果受害后,抗营养和抗消化酶类增加,营养质量下降(Haukioja,1990;康乐,1995;李镇宇等,1998)。最近的研究表明:球果的一种木蠹象害虫Pissodes strobi和外源MJA处理,都可诱导针叶树枝松脂(双萜)和挥发性单萜的从头合成,都可引起单萜合酶基因在转录水平的增加,但与MJA处理相比,虫害诱导的单萜合酶基因能快速表达,从而促进单萜挥发物的合成、积累和释放,表现为虫害诱导的反应更强(Fäldt et al., 2003;Martin et al., 2002;2003),与作者的结果一致。
油松球果小卷蛾的先期危害,使松果梢斑螟越冬幼虫趋向受害果取食(李新岗,2006),其结果造成2种害虫的食料竞争。在竞争中,小卷蛾幼虫大量死亡,而梢斑螟的死亡率却很低(李新岗, 2002)。但是,由于存活下来的小卷蛾幼虫提前脱果下地,使受害果内的梢斑螟幼虫寄生率提高(李宽胜,1992)。这说明趋向受害球果的梢斑螟,面临着天敌的威胁。
球果受害后,抗营养和抗消化酶类增加,营养质量下降,这与一般的诱导反应一致(娄永根等,1997;刘芳等,2004)。桦树(Betula sp.)叶片受到损伤时,能产生诱导抗虫性;而当幼嫩顶芽受到损伤时,却对植食性昆虫更敏感(Haukioja,1990)。油松针叶受害后,也能产生诱导抗虫性(李镇宇等,1998);但球果受到病虫危害或损伤后,则表现为对球果害虫的吸引(Fatzinger et al., 1985;Hanula et al., 1985;Valterova et al., 1995;Fidgen et al., 1996;Sadof et al., 1997;Kleinhentz et al., 1999)。甜菜夜蛾(Spodoptera exigua)幼虫危害小而幼嫩的马铃薯后对马铃薯叶甲(Leptinotarsa decemlineata)具有明显的引诱作用,但大而健康马铃薯则可直接引诱马铃薯叶甲(Bolter et al., 1997)。所以,可以推测幼嫩组织自身挥发成分的种类或数量不足以被某些害虫识别,只有受到危害或损伤后,才能被这些害虫识别,导致害虫危害,而引诱来的昆虫则有相应的适应机制,以应对幼嫩组织诱导后抗性成分增加、营养质量下降、天敌增加或竞争者存在造成的威胁。
桂连友, 刘树森, 陈宗懋. 2004. 外源茉莉酸和茉莉酸甲酯诱导植物抗虫作用及其机理. 昆虫学报, 47(4): 507-514. DOI:10.3321/j.issn:0454-6296.2004.04.016 |
桂连友, 陈宗懋, 刘树森. 2005. 外源茉莉酸甲酯处理茶树对茶尺蠖幼虫生长的影响. 中国农业科学, 38(2): 302-307. DOI:10.3321/j.issn:0578-1752.2005.02.014 |
康乐. 1995. 植物对昆虫的化学防御. 植物学通报, 12(4): 22-27. |
李宽胜. 1992. 油松球果害虫防治技术研究. 西安: 陕西省科学技术出版社.
|
李新岗. 2002. 油松球果害虫的危害机理研究. 西北农林科技大学学报, 30(2): 78-82. |
李新岗, 刘惠霞, 刘拉平, 等. 2006. 影响松果梢斑螟寄主选择的植物挥发物成分研究. 林业科学, 42(6): 71-78. |
李镇宇, 陈华盛, 袁小环, 等. 1998. 油松对赤松毛虫的诱导化学防御. 林业科学, 34(2): 43-50. DOI:10.3321/j.issn:1001-7488.1998.02.007 |
刘芳, 娄永根, 程家安. 2003. 虫害诱导的植物挥发物:植物与植食性昆虫及天敌相互作用的进化产物. 昆虫知识, 40(6): 481-486. DOI:10.3969/j.issn.0452-8255.2003.06.001 |
娄永根, 程家安. 1997. 植物诱导的抗虫性. 昆虫学报, 40(3): 320-331. DOI:10.3321/j.issn:0454-6296.1997.03.018 |
罗玉萍, 杨希楠, 姚容君, 等. 2001. 从松针中提取混合氨基酸方法研究. 天然产物与开发, 13(2): 41-42. |
姚延梼, 张淑改, 许茂红. 1998. 华北落叶松铜、钼含量及多酚氧化酶活性研究. 林业科学研究, 11(1): 94-98. |
Bolter C J, Dicke M, Van Loon J J A, et al. 1997. Attraction of colorado potato beetle to herbivore -damaged plants during herbivory and after its termination. J Cheml Ecol, 23(4): 1003-1023. DOI:10.1023/B:JOEC.0000006385.70652.5e |
Dicke M, Van Loon J J A. 2000. Multitrophic effects of herbivore-induced plant volatiles in an evolutionary context. Entomol Exp Appl, 97: 237-249. DOI:10.1046/j.1570-7458.2000.00736.x |
Dicke M. 2000. Chemical ecology of host-plant selection by herbivorous arthropods: a multitrophic perspective. Biochem System Ecol, 28: 601-617. DOI:10.1016/S0305-1978(99)00106-4 |
Dormont L, Roques A, Malosse C. 1997. Efficiency of spraying mountain pine cones with leoresin of swiss stone pine cones to prevent insect attack. J Chem Ecol, 23(10): 2261-2274. DOI:10.1023/B:JOEC.0000006672.05861.6c |
Dormont L, Roques A, Malosse C. 1998. Cone and foliage volatiles emitted by Pinus cembra and some related conifer species. Phytochemistry, 49(5): 1269-1277. DOI:10.1016/S0031-9422(98)00105-8 |
Fäldt J, Martin D, Miller B, et al. 2003. Traumatic resin defense in Norway spruce (Picea abies): Methyl jasmonate-induced terpene synthase gene expression, and cDNA cloning and functional characterization of (+)-3-carene sybthase. Plant Mol Biol, 51: 119-133. DOI:10.1023/A:1020714403780 |
Fatzinger C W, Merkel E P. 1985. Oviposition and feeding preferences of the southern pine coneworm for different host-plant materials and observations on monoterpenes as an oviposition stimulant. J Chem Ecol, 11(6): 689-699. DOI:10.1007/BF00988298 |
Fidgen L, Sweeney J. 1996. Fir coneworm, Dioryctria abietella, prefer cones previously exploited by spruce cone maggots. IUFRO. Working Party, Z7. 03-01, Cone and Seed Insects, 5-6
|
Hanula J L, Berisfori C W, DeBarr G L. 1985. Monoterpene oviposition stimulants of Dioryctria amatella in volatiles from fusiorm rust galls and second year loblolly pine cones. J Chem Ecol, 11(7): 943-952. DOI:10.1007/BF01012080 |
Heijari J, Nerg A, Kainulainen P, et al. 2005. Application of methyl jasmonate reduces growth but increases chemical defence and resistance against hylobius abietis in scots pine seedlings. Ent Exp Appl, 115(1): 117-124. DOI:10.1111/j.1570-7458.2005.00263.x |
Haukioja E. 1990. Induction of defenses in trees. Annu Rev Entomol, 36: 25-42. |
Kleinhentz M, Jactel H, Menassieu P. 1999. Terpene attractant candidates of Dioryctria sylvestrella in Maritime pine oleoresin, needles, liber and headspace samples. J Chem Ecol, 25(12): 2741-2756. DOI:10.1023/A:1020803608406 |
Kessler A, Baldwin I T. 2001. Defensive function of herbiovore-induced plant volatile emissions in nature. Science, 291(16): 2141-2144. |
Kessler A, Baldwin I T. 2002. Plant responses to insect herbiovory: The emerging molecular analysis. Annu Rev Plant Biol, 53: 299-328. DOI:10.1146/annurev.arplant.53.100301.135207 |
Martin D, Tholl D, Gershenzon J, et al. 2002. Methyl jasmonate induces traumatic resin ducts, terpenoid resin biosynthesis, and terpenoid accumulation in developing xylem of Norway spruce. Plant Physiol, 129(3): 1003-1018. DOI:10.1104/pp.011001 |
Martin D, Gershenzon J, Bohlmann J. 2003. Induction of volatile terpene biosynthesis and diurnal emission by methyl jasmonate in foliage of Norway spruce. Plant Physiol, 132(3): 1586-1599. DOI:10.1104/pp.103.021196 |
Sadof C S, Grant G G. 1997. Monoterpene composition of Pinus sylvestris varieties resistant and susceptible to Dioryctria zimmermani. J Chem Ecol, 23(8): 1917-1927. DOI:10.1023/B:JOEC.0000006479.39087.60 |
Thaler J S. 1999. Jasmonate-inducible plant defences cause increased parasitism of herbivores. Nature, 399(17): 686-688. |
Valterova L, Sjodin K, Vrkoc J, et al. 1995. Contents and enantiomeric compostion of monoterpene hydrocarbons in xylem oleoresin from four pinus species growing in Cuba. Comparsion of trees unattacked and attacked by Dioryctria horneana. Biochem Syst Ecol, 23(1): 115. |
Van Dam N M, Horn M, Mares M, et al. 2001. Ontogeny constrains systemic protease inhibeitor response in Nicotiana attenuata. J Chem Ecol, 22(3): 547-568. |
Zhu-Salzman K, Bi J, Liu T. 2005. Molecular strategies of plant defense and insect counter-defense. Insect Science, 12(1): 3-15. DOI:10.1111/j.1672-9609.2005.00002.x |