 2007, Vol. 43
2007, Vol. 43文章信息
- 史刚荣, 邢海涛.
- Shi Gangrong, Xing Haitao.
- 淮北相山8个树种叶片的生态解剖特征
- Eco-Anatomical Characteristics of Eight Tree Species in Xiangshan Mountain, Huaibei
- 林业科学, 2007, 43(3): 28-33.
- Scientia Silvae Sinicae, 2007, 43(3): 28-33.
-
文章历史
- 收稿日期:2005-12-29
- 修回日期:2006-05-24
-
作者相关文章


植被恢复是退化生态系统恢复重建的首要工作,也是我国恢复生态学的重点研究领域(彭少麟,1996)。生态系统的退化过程亦即生态系统的逆向演替过程,植物群落演替理论是指导退化生态系统恢复与重建的最重要的理论依据(Jordan et al., 1987)。植物的适应性与其生态分布、演替状况的关系是生态系统稳定性的基础。因此,如何从群落的层面上选择适宜的先锋植物种,建立生物学和生态学稳定的植物群落,一直影响着地带性植被恢复的进程(薛智德等,2004)。
结构是功能的基础,结构的差异和变化必然影响到生理生态功能,并进而影响到物种的分布。叶片是植物对环境变化最为敏感的器官之一,其形态和解剖结构特征被认为最能体现环境因子的影响及植物对环境的适应(Mott et al., 1982)。因此,研究叶片的结构及其对环境的响应,对阐释物种分布的格局具有重要意义。目前,对不同生境下同种植物叶形态解剖特征的比较研究不少(袁永明等, 1991;贺金生等,1994;费松林等, 1999;蔡永立等, 1999;2002;方精云等, 2000;Chabot et al., 1979;Mott et al., 1991;Rȏças et al., 1997;Bongers et al., 1990;Carpenter et al., 1975),对群落演替系列中优势种的生态解剖学也有研究(史刚荣等,2006;彭少麟等,2002;Strauss-Debenedetti et al., 1994),但通过比较退化生态系统恢复演替系列中主要种类叶片解剖特征的变化,探讨植物群落恢复演替的原因的研究却很少。本文在前期工作的基础上,以淮北相山混交林和杂灌丛的8个主要树种为研究对象,对其叶片的解剖特征及其可塑性进行了研究,以期为生态恢复不同演替阶段的树种选择和管理提供科学依据。
1 研究区概况研究地点为淮北相山,属暖温带半湿润季风气候,常年降水较少,年平均降水量约800~900 mm;年平均气温在14~17 ℃,全年无霜期220 d左右;日照时数2 300~2 500 h。土壤为石灰土及淋溶褐土。由于长期人为活动的影响,原有的暖温带落叶阔叶林已不复存在,大部分山丘为次生裸地,岩石裸露,蓄水能力极差。自然植被稀疏,主要植被类型有禾本科植物为优势种的草丛,酸枣(Ziziphus jujuba var. spinosa), 牡荆(Vitex negundo var. cannabifolia)为优势种的灌丛或灌草丛,以及局部发育良好的人工侧柏(Platycladus orientalis)林和混交林。
2 材料与方法 2.1 材料研究材料为相山东南坡混交林[郁闭度47%,侧柏和构树(Broussonetia papyrifera)、柘树(Cudrania tricuspidata)、野杏(Armeniaca vulgaris var. ansu)等树种占优势]和杂灌丛(全光照,酸枣、牡荆等树种占优势)群落中的8个主要树种(表 1)。取样时,每个群落每种植物随机选5个植株,按不同方向在树冠中部外表面取4片叶子,以消除方向不同引起的差异。沿新鲜叶片中脉两侧剪取长1 cm,宽0.5 cm的小片,置FAA固定液中固定和保存。
|
|

叶片结构采取常规石蜡切片法制片,切片厚度为12 μm,番红-固绿对染,加拿大树胶封片。叶表面结构用指甲油涂于叶片表面取印记,制成临时装片。在Motic Image Advanced 3.2软件下测量叶片厚度、上下表皮厚度、叶肉组织(包括栅栏组织和海绵组织)厚度、中脉厚度、维管束厚度、气孔密度、气孔长度等指标。每项指标测量40组数据。
2.3 统计分析叶片结构的可塑性指数(plasticity index,PI)参考Ashton等(1998)的计算公式:PI=1-x/X,式中:x为最小平均值,X为最大平均值。以主成分分析对叶片解剖性状进行因子分析。数据的统计分析在DPS数据处理系统(Data Processing System) 3.01下进行。
3 结果与分析 3.1 叶片的解剖特征8个树种叶片可分为2种类型:一类为异面叶,如柘树、扁担木、野杏、石榴和刺槐;另一类为全栅等面叶和异面叶混合型,如牡荆、酸枣、构树等,此类植物叶片在混交林中为异面叶,但在灌丛中却存在全栅等面叶。叶片厚度75.4~219.1 μm,以柘树最大,构树最小;上、下表皮均为单层细胞,上、下表皮厚度分别为12.8~36. 8 μm和5.6~20.0 μm,以酸枣和野杏最厚;上、下表皮角质膜厚度分别在1.3~4.0 μm和0.9~4.1 μ m,以酸枣和石榴最厚;除构树外,其余7个树种上表皮均无气孔分布,下表皮气孔密度90. 7~994.8 mm-2,以扁担木最少,牡荆最多。栅栏组织厚度35.0~113.9 μm,以酸枣和牡荆较厚,石榴最薄;海绵组织厚度8.9~99.9 μm, 以柘树最厚,构树最薄;栅栏组织海绵组织厚度比(S/P)为0.3~2.2,以构树最小,石榴最大(表 2)。
|
|
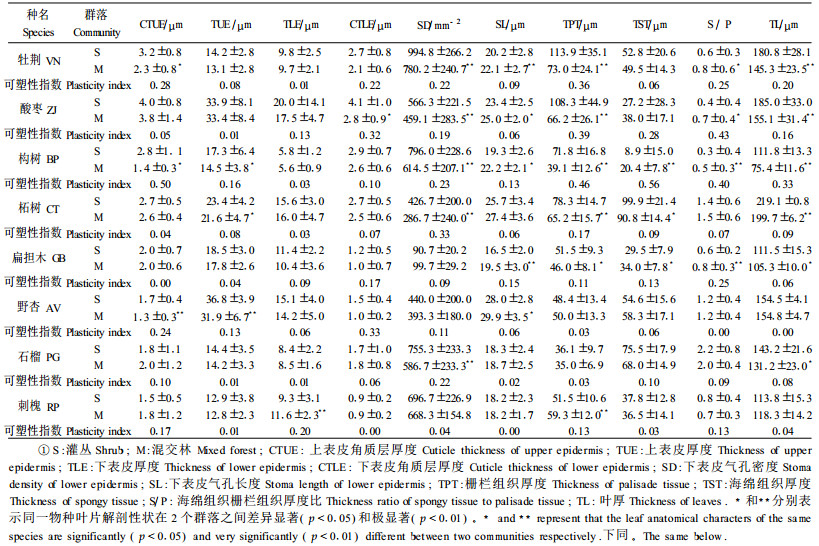
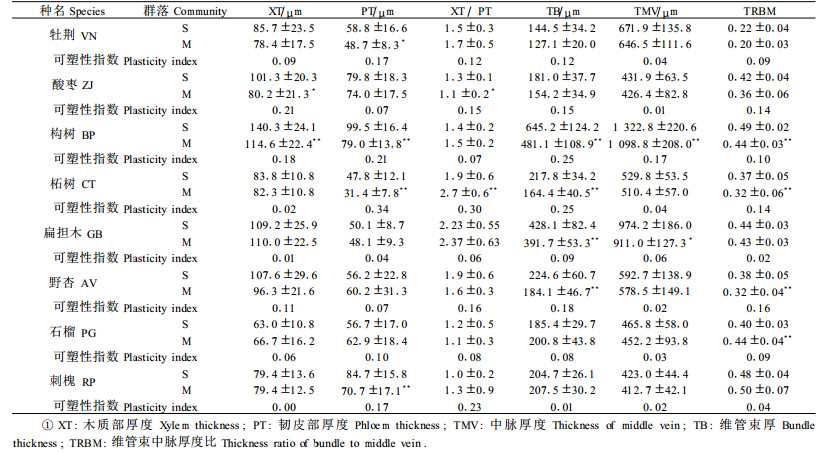
中脉以构树最为发达,而刺槐和酸枣较小。中脉厚度为412.7~1 3 22.8 μm,木质部厚度为63.0~140.3 μm,韧皮部厚度为31.4~99.5 μm,木质部韧皮部厚度比为1.0~2.37,维管束厚度为127.1~645.2 μm,维管束中脉厚度比为0.20 ~0.50(表 3)。
|
|
不同树种叶片对不同群落环境表现出的可塑性差异很大,其可塑性指数大小依次为:构树>酸枣>牡荆>柘树>野杏>扁担木>刺槐>石榴(图 1a)。叶片的不同解剖性状对不同群落环境的可塑性也有很大的差异,其中以栅栏组织厚度、S/P、气孔密度、表皮角质层厚度、海绵组织厚度、韧皮部厚度、木质部韧皮部厚度比、维管束厚度以及叶片厚度等性状可塑性较大(图 1b),说明这些性状易受环境条件的影响。叶片解剖可塑性与各树种的生活型关系不大(图 1d),但与它们在群落中的生态位密切相关,与优势种相比,亚优势种叶片解剖可塑性显著较小,广布优势种和局部优势种之间在叶片解剖可塑性上差异不显著(图 1c)。

|
图 1 不同树种叶片解剖性状的可塑性比较 Fig. 1 Comparison on plasticity of leaf anatomical characters of different species WD: 广布优势种 Widely dominant species; LD: 局部优势种 Locally dominant species; SUD: 亚优势种 Subdominant species. |
由表 2可见,灌丛群落植物叶片表现出阳生特点:厚的表皮及角质层,厚的叶片和栅栏组织,小的S/P,小而多的气孔,发达的输导组织。与灌丛群落相比,混交林植物叶片结构却不同程度地表现出阴生特点:表皮及角质层薄,叶片较薄,栅栏组织不发达,大的S/P,气孔大而少,输导组织不发达。
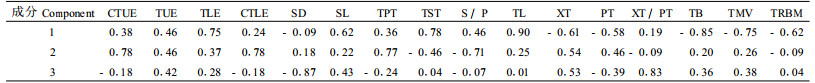
3.3 叶片解剖性状的多元统计分析对16个叶片解剖性状进行主成分分析(前三维主成分反映原始数据信息的74.31%)。结果表明,叶片厚度、维管束厚度、海绵组织厚度、上下表皮角质层厚度、栅栏组织厚度、气孔密度、木质部韧皮部厚度比等性状对主成分的贡献较大(表 4)。为了与一般解剖性状相区别,将这些在不同物种之间或同一物种不同生境之间存在较大变异的性状称为关键解剖性状(key anatomical characters)。根据这些变量的原始含义可以指出3个主成分功能含义:第1主成分(Y1)综合反映了植物叶片对光照强度和水分的适应(占总信息量的34.59%),其中作用较大的性状包括叶片厚度、维管束厚度、海绵组织厚度;第2主成分(Y2)主要表现植物叶片对光照强度的适应(占总信息量的22.69%),其中以上下表皮角质层厚度、栅栏组织厚度等性状的贡献较大;第3主成分(Y3)主要表现植物叶片对水分的适应(占总信息量的17.04%),其中以气孔密度、木质部韧皮部厚度比等的贡献较大。
|
|
以第1主成分和第2主成分为轴,对混交林和杂灌丛中的8个树种进行排序(图 2),排序结果表明,8个主要树种大致可分为3类:Ⅰ类为阳性旱生植物,包括酸枣、牡荆、柘树和野杏;Ⅱ类耐荫性中生植物,包括刺槐、扁担木和石榴;Ⅲ类为阳性中生植物,仅构树1种。从图 2中可以看出,同一物种因其所在的群落不同而在排序轴上具有一定的距离,距离的大小因物种的不同而不同,总体上看,阳性树种(如酸枣、牡荆、柘树、野杏和构树)大于耐荫性树种(如刺槐、扁担木和石榴),可见,阳性树种比耐荫性树种在叶片结构上具有较大的可塑性,这与图 1a中的结果相一致。尽管如此,但不同群落的同一物种大都聚在同一大类之中,说明叶片结构受着遗传和环境因素的双重影响。

|
图 2 8个树种叶片解剖性状的主因子排序(占总信息量的57.27%) Fig. 2 Principle component analysis ordination on leaf anatomical characters of eight species 1. 灌丛 Shrub; 2. 混交林 Mixed forest. |
关于不同演替系列中植物的可塑性一直有不同的结论。很早就有人认为,早期演替植物种比晚期演替植物种有更大的适应能力(Bazzaz, 1979)。Zangerl等(1983)认为后期种对光和温度缺少可塑性,这一观点得到了Raich(1989)的试验支持。对温带森林的研究表明,中间演替树种比早期和晚期演替树种有更大的表型可塑性(Neuner et al., 1995),中间种能够适应较宽范围的生态环境(Abrams et al., 1995)。本研究的群落均处于演替早期阶段,结果显示,叶片解剖可塑性在不同树种之间差异很大,各树种叶片的解剖可塑性与其生活型关系不大,却与植物在群落中的生态位有关。比较而言,优势树种(如构树、酸枣、牡荆、柘树等)大于亚优势树种(如野杏、刺槐、石榴)。
叶片的不同解剖学性状的可塑性大小存在很大差异。可塑性较大的性状包括栅栏组织厚度、海绵组织栅栏组织厚度比、气孔密度、表皮角质层厚度、海绵组织厚度、韧皮部厚度、木质部韧皮部厚度比、维管束厚度以及叶片厚度等(图 1b)。主成分分析表明,叶片厚度、维管束厚度、海绵组织厚度、气孔密度、木质部韧皮部厚度比、栅栏组织厚度和S/P等性状,在物种之间或同一物种不同生境之间存在较大变异,称为关键解剖性状。由于这些性状更能反映环境对叶片解剖结构的影响,因而在生态解剖学研究中具有重要作用。
表型可塑性使得物种具有更宽的生态幅和更好的耐受性,可以占据更加广阔的地理范围和更加多样化的生境,即成为生态位理论中的广幅种(Sultan,1995)。优势种(特别是广布优势种)由于在时间和空间上需要经历或占据不同的群落环境,因而面临不同的环境压力。由于叶片结构的可塑性,同一树种叶片在不同演替群落中表现不同的结构特点。在灌丛群落中,植物叶片表现出阳生特点:厚的表皮及角质层,厚的叶片和栅栏组织,小的S/P,小而多的气孔,发达的输导组织。在混交林中,却表现出不同程度的阴生特点:表皮及角质层薄,叶片和栅栏组织较薄,S/P大,气孔大而少,输导组织不发达。正是叶片结构的这种可塑性,使得它们既能适应恢复演替早期的干旱生境,又能忍受演替后期的荫蔽条件,从而成为优势种。
4.2 叶片的生态类型及其群落学意义不同物种长期生长于特定的生境条件下,必然形成一些相应的适应特征,而植物的适应特征又反过来影响植物的分布。本文研究结果表明,8个树种叶片结构对其生境表现出多样化的适应方式,大致可分为3类,即阳性旱生植物、耐荫性中生植物和阳性中生植物。尽管酸枣、牡荆和柘树3个广布优势种的叶片均具有较大的表型可塑性,但在2个群落中均表现出阳性旱生植物的特征,这些特征与恢复演替早期较强的光照和干旱条件相适应,可认为是其成为广布优势种的解剖学基础。2个局部优势种均为中生植物,其中,作为乔木层优势种的构树为阳性中生植物,灌木层优势种之一的扁担木为耐荫性中生植物。显然,在淮北相山,水分是限制这2个局部优势树种在群落中水平分布的重要因子,而光照则是限制植物在层片中垂直分布的主导因子。
演替早期植物或先锋植物被认为具有许多阳生性植物的特征,演替后期或顶极群落植物则是对荫蔽条件具有较强忍受力的类群(赵平等, 2003)。杂灌丛中的3个优势种(酸枣、牡荆、柘树)均为阳性树种,而混交林中的5个优势种中,只有1个耐荫性植物(扁担木)。这一结果表明,演替早期植被中优势种多为阳性植物,随着演替的进展,耐荫性植物将不断加入到优势种的行列。
可见,相山恢复演替早期植被的优势种的叶片都表现出一定的解剖可塑性,这是植物适应其异质生境的重要机制。叶片的解剖特征不仅是植物对环境条件的直接效应的反映,而且是植物在群落中空间分布及其群落性质的反映。运用多元分析方法,通过比较先锋群落到顶极群落一些主要种类叶片的解剖学特征的差异,将有助于理解植物在群落中的作用,进而揭示植物群落演替的原因。
蔡永立, 达良俊. 2002. 亚热带东部壳斗科6种常绿植物叶的生态解剖. 应用与环境生物学报, 8(5): 460-466. DOI:10.3321/j.issn:1006-687X.2002.05.003 |
蔡永立, 王希华, 宋永昌. 1999. 中国东部亚热带青冈种群叶片的生态解剖. 生态学报, 19(6): 844-849. DOI:10.3321/j.issn:1000-0933.1999.06.013 |
方精云, 费松林, 樊拥军, 等. 2000. 贵州梵净山亮叶水青冈解剖特征的生态格局及主导因子分析. 植物学报, 42(6): 636-642. DOI:10.3321/j.issn:1672-9072.2000.06.016 |
费松林, 方精云, 樊拥军, 等. 1999. 贵州梵净山亮叶水青冈叶片和木材的解剖学特征及其与生态因子的关系. 植物学报, 41(9): 1002-1009. DOI:10.3321/j.issn:1672-9072.1999.09.018 |
贺金生, 陈伟烈, 王勋陵. 1994. 高山栎叶的形态结构及其与生态环境的关系. 植物生态学报, 18(3): 219-227. DOI:10.3321/j.issn:1005-264X.1994.03.010 |
彭少麟. 1996. 恢复生态学与植被重建. 生态科学, 15(2): 26-31. |
彭少麟, 李跃林, 余华, 等. 2002. 鼎湖山森林群落不同演替阶段优势种叶生态解剖特征研究. 热带亚热带植物学报, 10(1): 1-8. DOI:10.3969/j.issn.1005-3395.2002.01.001 |
史刚荣, 汤盈, 张铮. 2006. 淮北相山恢复演替群落优势树种叶片的生态解剖. 植物生态学报, 30(2): 314-322. DOI:10.3321/j.issn:1005-264X.2006.02.015 |
薛智德, 韩蕊莲, 侯庆春, 等. 2004. 延安地区5种灌木叶旱性结构的解剖研究. 西北植物学报, 24(7): 1200-1206. DOI:10.3321/j.issn:1000-4025.2004.07.008 |
袁永明, 彭泽祥, 陈家瑞. 1991. 黄华族(豆科)叶的解剖学性状的系统学和生态学意义. 植物学报, 33(11): 840-847. |
赵平, 曾小平, 彭少麟. 2003. 植被恢复树种在不同实验光环境下叶片气体交换的生态适应特点. 生态学杂志, 22(3): 1-8. DOI:10.3321/j.issn:1000-4890.2003.03.001 |
Abrams M D, Mostoller S A. 1995. Gas exchange, leaf structure and nitrogen in contrasting successional tree species growing in open and understorey sites during a drought. Tree Physiol, 15: 361-370. DOI:10.1093/treephys/15.6.361 |
Ashton P M S, Olander L P, Berlyn G P, et al. 1998. Changes in leaf structure in relation to crown position and tree size of Betula papyrifera within fire-origin stands of interior cedar-hemlock. Can J Bot, 76: 1180-1187. |
Bazzaz F A. 1979. The physiological ecology of plant. Ann Rev Ecol Syst, 10: 351-371. DOI:10.1146/annurev.es.10.110179.002031 |
Bongers F, Popma J. 1990. Leaf characteristics of the tropical forest flora of Los Tuxtlas, Mexico. Bot Gaz, 151: 354-365. DOI:10.1086/337836 |
Carpenter S B, Smith N B. 1975. Stomatal distribution and size in southern Appalachian hard woods. Can J Bot, 53: 1153-1156. DOI:10.1139/b75-137 |
Chabot B F, Jurik T W, Chabot J F. 1979. Influence of instantaneous and integrated light-flux density on leaf anatomy and photosynthesis. Amer J Bot, 66: 940-945. DOI:10.1002/j.1537-2197.1979.tb06304.x |
Jordan W R, Gilpin M E, Aber J D. 1987. Restoration ecology: A synthetic approach to ecological research. Cambridge: Cambridge University Press.
|
Mott K A, Gibson A G, O'Leary J W. 1982. The adaptive significance of amphistomatic leaves. Plant Cell Environ, 9: 455-460. |
Mott K A, Michaelson O. 1991. Amphistomy as an adaptation to high light intensity in Ambrosia cordifolia (Compositae). Amer J Bot, 78: 76-79. DOI:10.1002/j.1537-2197.1991.tb12573.x |
Neuner G, Bannister P. 1995. Frost resistance and susceptibility to ice formation during natural hardening in relation to leaf anatomy in three evergreen tree species from New Zealand. Tree Physiol, 15: 371-377. DOI:10.1093/treephys/15.6.371 |
Raich J W. 1989. Seasonal and spatial variation in the light environment in the tropical Dipterocarp forest and gaps. Biotropica, 21: 229-302. |
Rȏças G, Barros C F, Scarano F R. 1997. Leaf anatomy plasticity of Alchornea triplinervia (Euphorbiaceae) under distinct light regimes in a Brazilian montane Atlantic rain forest. Trees, 11: 469-473. |
Strauss-Debenedetti S, Berlyn G P. 1994. Leaf anatomical responses to light in five tropical Moraceae of different successional status. Am J Bot, 81: 1582-1591. DOI:10.1002/j.1537-2197.1994.tb11470.x |
Sultan S E. 1995. Phenotypic plasticity and plant adaptation. Acta Botanica Neerlandica, 44: 363-383. DOI:10.1111/j.1438-8677.1995.tb00793.x |
Zangerl A R, Bazzaz F A. 1983. Responses of early and late successional species of Polygonum to variations in resource availability. Oecologia, 56: 397-404. DOI:10.1007/BF00379719 |