 2007, Vol. 43
2007, Vol. 43文章信息
- 郑勇平, 孙鸿有, 董汝湘, 华朝晖, 唐淑琴, 张建章, 傅顺华.
- Zheng Yongping, Sun Hongyou, Dong Ruxiang, Hua Zhaohui, Tang Shuqin, Zhang Jianzhang, Fu Shunhua.
- 杉木不同世代不同类型种子园遗传改良增益研究
- A Study on Realized and Genetic Gains of Different Generations and Types in Seed Orchards of Chinese Fir (Cunninghamia lanceolata)
- 林业科学, 2007, 43(3): 20-27.
- Scientia Silvae Sinicae, 2007, 43(3): 20-27.
-
文章历史
- 收稿日期:2006-02-05
-
作者相关文章



2. 浙江林学院林业与生物技术学院 临安 311300;
3. 浙江省林业种苗管理总站 杭州 310020;
4. 浙江遂昌县林业局 遂昌323300;
5. 浙江淳安县姥山林场 淳安 311700;
6. 浙江龙泉市林业局 龙泉 323700
2. School of Forestry and Biotechnology, Zhejiang Forestry College Lin'an 311300;
3. Forestry Seed and Stock Controlling Station of Zhejiang Province Hangzhou 310020;
4. Forestry Bureau of Suichang County Suichang 323300;
5. Laoshan Forest Farm of Chun'an County Chun'an 311700;
6. Forestry Bureau of Longquan County Longquan 323700
国内外林木群体良种选育,基本上是沿着选优、建园、子代测验、再选择、再建园……的轮回选育路线,建立一代又一代种子园,一步步从低世代种子园向高世代种子园发展。国外的种子园,已发展到第3代(沈熙环,1994)。国内以杉木(Cunninghamia lanceolata)种子园发展最快,已从1代、1.5代发展到2代种子园,现已开始建立2.5代种子园,并且已出现第3代育种群体。国内的种子园,基本上是育种群体与生产群体合二为一(王明庥,2001),每个种子园既是一代轮回选择的结果,又是新一代轮回选择的基础,因此,从总体上全面研究各世代、类型种子园遗传改良进展,对探讨杉木及其他用材树种群体良种选育策略具有重要意义。
关于种子园遗传改良进展,国内外先后有较多报道,但是,大多数是某一个或少数几个种子园遗传增益试验的结果(李锦清等,1991;吴隆高等,1993a; 1993b;陈代喜等,1999;邹斌等,2003)。由于各自的试验条件、试验方法和试验对照等的不同,其报道的遗传增益相互之间缺乏可比性,也难以正确看出世代间的发展趋势,迄今未见将各个世代多个种子园子代集中在同一个试验中,从世代的横向和纵向空间上对种子园遗传增益进行系统的研究报道。国内,徐金良等(1997)曾对杉木1代种子园、1.5代种子园、种源种子园等作过比较试验,但也只是涉及到5个种子园的1地1年的试验。国外,Shelbourne(1991)以辐射松(Pinus radiata)遗传信息为例,对可能组成的10种种子园的子代生长增益进行过预测,但只是理论上的推算,未经过试验验证。一些试验表明,理论预测值与试验结果,往往有较大差异(Squillace, 1989;Wright et al., 1996)。
有鉴于此,本项研究将杉木现有的1代种子园、1.5代种子园、2代种子园、优良种源种子园和优良双系种子园5种世代、类型种子园中的17个种子园子代,集中在同一个试验中进行子代测验,研究其遗传增益及其变化趋势,进而探讨杉木和其他用材树种群体良种选育的持续高效发展策略。本文主要是从种子园的世代、类型整体上,进行研究分析。
1 材料与方法 1.1 参试种子园和无性系组成为保证整体试验的准确性,参试的种子园都是在国家林业局和浙江省联营的7个林木良种基地内建立的,在非林木良种基地建立的种子园均未参试。共有5种世代、类型17个种子园参试,其中,1代、1.5代和2代种子园各5个,优良种源种子园和优良双系种子园各1个(唯一的)。参试的15个1代、1.5代和2代种子园,其中有1个种子园无性系全部来自浙江省(本地)种源,其他14个种子园,其无性系组成基本上是以浙江省种源为主,同时吸纳杉木中心产区广西、贵州、湖南、福建等省(区)种源的部分无性系材料(表 1)。优良种源种子园是由广西柳州地区优良种源的优树无性系组成,其中融水种源的无性系占69.8%。优良双系种子园是由全同胞子代测定中选出的最优组合的2个亲本无性系组成,该双系也是浙江省首批审定的2个优良单亲本家系的亲本。
|
|

按照中华人民共和国《林木种子检验方法》国家标准的取样法GB2772-81(国家标准总局,1982),各个种子园在种子全部采集处理后,从种子堆不同部位等量取种,充分混合后,用四分法提取试验种子。试验“对照”的种子,为浙南庆元县、浙中淳安县和浙北临安市三地未经遗传改良的普通商品种子或一般林分的种子(简称对照或普种)。试验种子经过品质检验后,进行育苗、造林试验。
1.3 试验林造林设计试验地设在浙江省杉木主产区临安市、淳安县、遂昌县和龙泉市4个地点。连续进行2年(次)试验。第1年(次)试验,1996年采种,1997年育苗,1998年造林;第2年(次)试验,1997年采种,1998年育苗,1999年造林。采用完全随机区组设计,5个区组,6株(第1次试验)或8株(第2次试验)小区。株行距2 m×2 m。造林后第2年春天,对试验林内死缺植株用保护行中同一种子园植株带土补植或用1年生大苗补植。
1.4 生长调查测定自造林后第2年起,每年逐株调查测定树高、胸径、冠幅、病虫害等。最后1次测定(2004年),在临安试验点增加侧枝情况调查,定位测定主干离地面1.5~3.5 m区段内的侧枝盘数(即节数)、盘间(节间)长度、每盘(节)一级侧枝数、一级侧枝密度(即主干平均1 m长度内的一级侧枝数)等。试验林每个植株都有固定编号,每次调查时,逐株对号记录。
1.5 统计分析原始数据以小区平均数为基数,采用DPS2000软件系统进行多年多地点多重复方差分析,其线性数学模式为Xijkl=μ+gi+yj+sk+bl(jk)+(gy)ij+(gs)ik+(gys)ijk+(ys)jk+eijkl,式中gi为种子园效应;yj为试验年份(次)效应;sk为试验地点效应;bl(jk)为年份(次)内地点内区组效应;(gy)ij为种子园×试验年份(次)效应;(gs)ik为种子园×试验地点效应;(gys)ijk为种子园×试验年份(次)×试验地点效应;(ys)jk为年份(次)×地点效应;eijkl为试验误差。
多重比较采用Duncan氏新复全距法。秩相关采用Spearmen模式。种子园遗传率估算采用公式
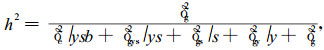
|
式中δg2为种子园群体遗传方差分量;δgy2、δgs2、δgys2和δe2分别为种子园×试验年份(次)、种子园×试验地点、种子园×试验年份(次)×试验地点、试验误差的方差分量。
2 结果与分析 2.1 种子园子代生长性状变异分析方差分析(表 2)表明:种子园之间在子代树高、胸径、材积上的差异都达到极显著水平,但是,种子园×试验年份(次)、种子园×试验地点、种子园×试验年份(次)×试验地点的F值都未达显著水平,其值大都小于1.0。这说明种子园之间在子代生长上的差异是相当稳定的,其在不同年份(次)试验、不同试验地点的生长大小排序,没有显著变化。这个结果反映出同一种子园在不同年份结实的种子,尽管由于不同年份环境条件、随机交配等因素变化的影响,而使其遗传组成有所不同,但是如无特殊因素影响,这种差异并未达显著水平。这个结果也反映出,种子园由于是多个无性系组成的群体,其子代对环境条件的适应性相对较强,与环境互作不明显,因而在不同试验地点生长大小排序变化也不显著。
|
|
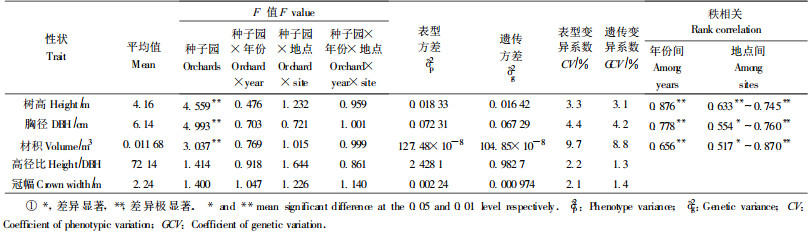
对试验林年份(次)间、地点间的秩相关分析,也验证了上述试验结果,秩相关系数都达到显著或极显著水平。从变异系数来看,树高、胸径、材积的遗传变异系数与表型变异系数相差很小,表明在本试验结果中,种子园在子代生长上的差异,基本上是遗传变异引起的。
2.2 不同世代、类型种子园子代生长增益总体分析参试的17个种子园,子代生长都优于对照,其中,树高、胸径、材积生长大于对照达到显著水平的种子园数,分别占82.4%、88.2%和76.5%。从种子园世代、类型总体上看,5种不同世代、类型种子园之间,子代生长增益不仅有较大差异,而且表现出明显的变化趋势(表 3)。
|
|
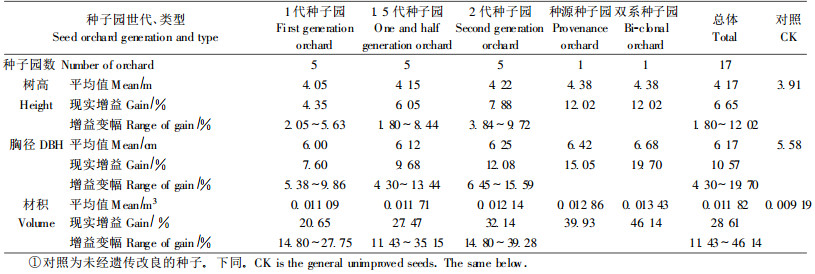
种子园子代平均树高比对照大6.65%,最大达12.02%,最小仅1.80%。从种子园世代、类型看,1代、1.5代和2代种子园,种源种子园,双系种子园分别比对照增长4.35%、6.05%、7.88%、12.02%和12.02%。
2.2.2 胸径种子园子代胸径生长的增幅比树高增幅大,其平均增幅为10.57%,变动于4.3 0%~19.70%。1代、1.5代和2代种子园,种源种子园,双系种子园分别比对照大7.60%、9.68%、12.08%、15.05%和19.70%。
2.2.3 材积种子园子代平均材积比对照大28.61%,变动于11.43% ~46.14%。1代、1.5代、2代种子园,以及种源种子园,双系种子园,分别比对照大20.65%、27.47%、32.14%、39.93%和46.14%。
所以,从整体上看,以优良双系种子园子代增益最大,优良种源种子园次之,二者在材积生长上的增益分别为2代种子园平均增益1.44倍和1.24倍。在3种世代种子园间,1代、1.5代和2代种子园,子代树高、胸径、材积的生长增益都是逐级递增的,表明每种世代种子园都取得了较好的改良效果。
从单个种子园多重比较来看,参试的17个种子园,也是以双系种子园增益最大,种源种子园次之,其后是2代种子园中最优种子园。双系种子园子代材积增益是最优的2代种子园增益的1.17倍,优良种源种子园与最优的2代种子园基本持平,为其1.02倍。
其实,双系种子园,在种子品质上也表现优良,除涩籽率改良增益幅度略小外,种子生活力和活力都名列前茅(孙鸿有等,2003)。双系种子园之所以有如此优良的表现,主要是因为该双系是在双亲本子代测验中选出的最优组合,它们本身也是浙江省首批审定的2个优良家系的亲本,具有优良的遗传基础。
同样,优良种源种子园子代的良好表现也是有其优良遗传基础的。据全国杉木种源试验协作组(1994)种源试验结果的评定和浙江省林木良种审定委员会的审定,广西融水、贵州锦屏、福建南平等6个种源为适宜浙江省造林的高产类型优良种源,其中,融水种源被评为全国极高产类型。本试验中的优良种源种子园是由广西柳州地区优良种源的优树无性系组成,其中,该区融水种源占69.8%。参试的1代、1.5代和2代种子园,其无性系组成以浙江种源为主,虽然其中也包含有优良种源的优树、优良家系的材料,但仅仅是一部分,其中融水种源占的比例更少,其子代增益当然难以胜过优良种源种子园。
2.3 3种世代种子园子代生长增益变化趋势分析在1代、1.5代、2代种子园中,参试的各个种子园虽然子代生长都大于对照,但是Duncan氏新复全距检验表明,三者都有少数种子园与对照的子代生长差异未达显著水平,其中在材积生长上,未达显著水平的种子园分别占40%、20%和20%。这充分证明种子园子代遗传测验对种子园建设的重要性,在生产上,对于一个新建的世代较高的种子园,在未经子代测验之前,不要轻易地以它取代世代较低的种子园。
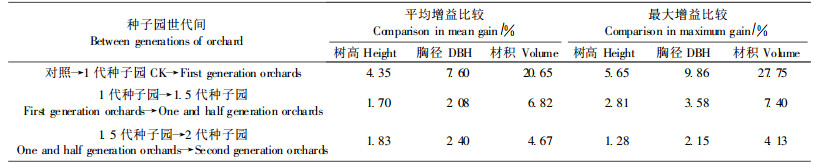
1代、1.5代和2代种子园之间,子代生长平均增益虽然是逐级递增的,但是,增幅却逐级快速下降。以“对照”为统一计算基础的子代材积生长增益,从对照→1代种子园→1. 5代种子园→2代种子园,世代之间的增益由1代的20.65%下降到1.5代的6.82%和2代的4. 63%。这是每种世代多个种子园相对于“对照”的平均增益变化趋势的反映。为避免增益过小的种子园对研判不同世代种子园之间增益变化趋势的影响,试验又以3种世代种子园中最优种子园的增益进行比较,结果同样表现出快速下降的趋势,与对照相比较,不同世代之间材积增益幅度由27.75%下降到7.28%和4.13%(表 4)。这种快速下降趋势的产生,可能一是由于1代种子园建设起点较高,吸收了中心产区部分优树无性系,因而增益相对较高;二是由于2代种子园建园时间较晚,参试时,正处于结实投产初期,对其子代生长增益会有一些影响,等等。除此之外,这种下降趋势,也意味着我国杉木种子园建设在向高世代种子园过渡中,可能存在一些需要改进的潜在问题。
|
|
种子园之间子代高径比的差异未达显著水平(表 2),但测定结果仍反映出一定的变化趋势,平均比对照小2.65%,其中1代、1.5代和2代种子园,种源种子园,双系种子园分别比对照小1.18%、2.05%、3.78%、3.65%和7.57%。这表明,种子园子代在幼林阶段,茎干削度要比对照略大些,并且随着生长增幅的增加而有所加大,这主要是由于种子园子代胸径增幅大于树高所致(表 5)。
|
|
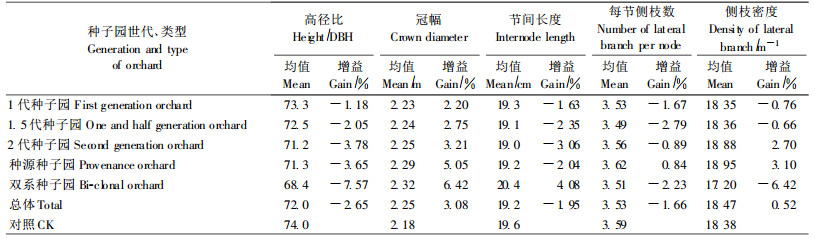
冠幅是杉木优树选择和家系选择的一个指标,一般选择窄冠型。试验结果表明,种子园子代冠幅平均比对照大3.08%,并且随着茎干生长增幅的加大而相应增大,但并非同比增大,其平均增幅仅为树高、胸径增幅的50%和30%(表 5)。这说明相对于树高、胸径生长增幅而言,对冠幅的选择还是有效果的。
2.4.3 侧枝性状侧枝性状也是杉木优树选择和家系选择的指标之一,一般选择枝叶浓密型。试验结果表明,种子园在子代主干节间长度上的差异达到显著水平,F值2.385**,每节一级侧枝数、一级侧枝密度的差异未达显著水平,F值分别为0.663和1.551。其中,由多系组成的1代、1.5代、2代种子园和种源种子园,子代节间长度平均比对照小1.63% ~ 3.06%,而在每节侧枝数和侧枝密度上无一定变化规律,二者增幅分别变动于-2.79% ~ 0.84%和-0.66% ~ 3.10%之间。双系种子园子代节间长度比对照大4.08%,每节侧枝数和侧枝密度则分别比对照小2.23%和6.42%。显然,多系种子园的遗传改良对于子代侧枝性状基本上没有影响,而双系种子园则有一定的影响,这与双系种子园的无性系分枝特性有一定关系(表 5)。
2.5 不同世代、类型种子园子代生长遗传增益分析 2.5.1 遗传率的估算遗传学的理论认为,决定个体性状遗传的是基因,而决定群体性状遗传的是基因频率,不同的种子园,由于无性系组成不同,基因频率必然不同,这必然影响种子园的子代生长。因此有必要通过方差分析,将试验中的种子园遗传方差从表型方差中分剖出来,估算其所占的比率(遗传率)(马育华,1982;王明庥,2001),从而分析出本试验中的种子园子代生长的遗传增益。以种子园群体平均值为基础,估算了各性状的遗传率。结果表明(表 6),种子园群体的性状遗传率估值,以树高、胸径和材积最大,分别为0.896、0.930和0.822;其后依次为冠幅、高径比、主干节间长度和一级侧枝密度,主干每节侧枝数为0。这表明,在本试验中,种子园之间子代树高、胸径、材积生长上的差异,主要是由于群体遗传基础差异造成的。
|
|
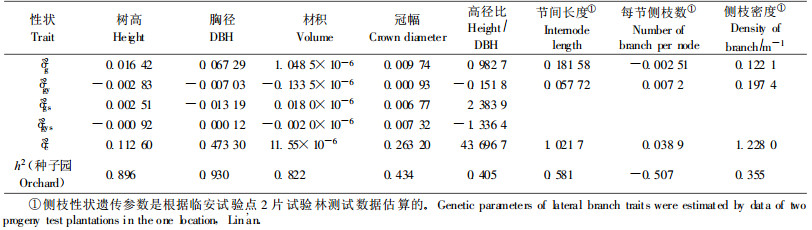
以种子园群体遗传率的估值,估测了种子园子代生长遗传增益。其遗传增益大小,取决于其相对于对照的增幅大小和群体性状遗传率大小。估算结果表明,17个种子园子代树高、胸径、材积平均遗传增益分别为6.08%、9.93%和23.56%。其中1代、1.5代和2代种子园,优良种源种子园,优良双系种子园,子代树高遗传增益依次为3.90%、5.41 %、7.06%、10.77%和10.77%;胸径遗传增益依次为7.07%、9.00%、11.23%、14.00%和18.32%;材积遗传增益为16.97%、22.58%、26.42%、32.82%和37.93%。
在3种世代种子园中,增益最小的种子园,其树高、胸径、材积的遗传增益只有1.61 %、4.00%和9.40%,而增益最大的种子园分别达到8.71%、14.50%和32.29%(表 7)。
|
|
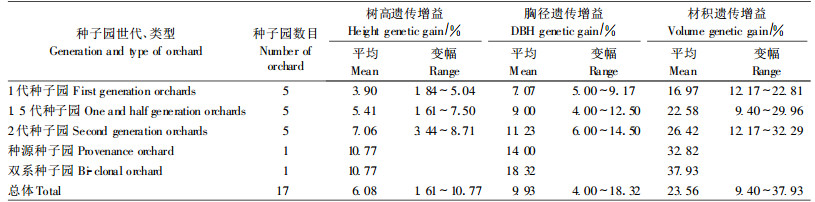
优良双系种子园表现最佳,子代生长优于参试的各个种子园,其材积现实增益高达4 6.14%,遗传增益37.93%,为1代、1.5代和2代种子园,种源种子园增益的2.23、1.68、1.44和1.15倍。看来人们担心的双系种子园因自交机会增多会对后代产生的不利影响,在杉木来说,实际上是不大的,在自由授粉的情况下,由于生物有避免自交的机制,其自交机会可能是很小的。本试验中双系种子园这样大的增益,虽然只是来自一个种子园,但是,各地在子代测验和双列交配试验中,已经选出一批增益远大于此双系种子园的优良家系与优良组合(方乐金等,1998;王欣,1991;阮梓材等,1998;李锦清等,1990;李寿茂等,1999;陈代喜等,1997;张建章等,1991;张建忠等,2005;洪昌端等,1992;赵剑斌等,1996;王赵民等,1988),不仅在一定程度上验证了双系种子园的子代生长优势,证明双系种子园的巨大优越性和发展潜力,也为发展双系种子园提供了优良的种质资源基础。今后的种子园建设,应将双系种子园列为重点之一。北美在栎树(Quercus)、瑞典在桦木(Betula)等树种的育种计划中已将双系种子园列入其中(Steiner,1993;Werner, 1993)。面对我国林业发展的新形势,建立双系种子园,实现工业原料林用种的品种化,应是今后的一个重要发展方向。
优良种源种子园表现突出,其材积现实增益和遗传增益为39.93%和32.82%,不仅明显高于2代种子园的平均增益,甚至与2代种子园中最优种子园的增益基本持平。此优良种源种子园是用广西柳州地区优良种源中的优树无性系建成的,实际上是优良种源区的1代种子园,显然,它包含了优良种源和优树双重增益。它的突出表现,显示出优良种源中优质种质资源潜在的遗传增产优势,在林木长期育种和高世代种子园建设中,应充分发掘并优先利用。
1代、1.5代和2代3种世代种子园增益逐级增长,其子代材积现实增益分别为20.65%、27.47%和32.14%,遗传增益为16.97%、22.58%和26.42%。这是各世代多个种子园在同一个多次多点试验中,相对于未经遗传改良的普通种子的平均增益,虽然各种子园遗传基础有所不同,但仍在一定程度上反映了杉木3种世代种子园的遗传改良增益水平。国外,Hodge等(1989)曾总结报道美国湿地松(Pinus elliottii)未去劣1代种子园、去劣1代种子园和改良1代种子园,15年生单株材积分别比普种大10%、15%和19%。Lee (1994)报道新西兰辐射松种子园现阶段材积遗传增益为15%~20%。Matziris (2000;2005)报道希腊阿勒颇松(Pinus halepensis)、欧洲黑松(Pinus nigra)未去劣1代种子园材积增益分别为13.0%和24.0%,去劣1代种子园分别为21.5%和32%。周显昌等(1999)报道,加拿大白云杉(Picea glauca)1.5代种子园遗传增益为10%~1 5%,小干松(Pinus contorta)为16%。Rosvall等(2004)报道,瑞典种子园现阶段材积增益为25%,到2010—2020年增益可达35%。虽然本文结果不能直接与这些报道相比较,但从中可以折射出,我国杉木3种世代种子园建设基本上是成功的,增益是大的。
3种世代种子园子代生长平均增益,虽然是逐级增长的,但世代层次之间的增幅却呈现明显下降趋势,其材积的现实增益,由未经遗传改良的普种→1代种子园→1.5代种子园→ 2代种子园,世代层次间增幅由1代种子园的20.65%下降为1.5代的6.82%和2代的4.67%,降幅很大。国外同类种子园也有这种情况,但降幅小得多。这种下降趋势并非不可避免,瑞典在长期育种计划中,提出小干松第1代改良获得增益6% ~8%,以后每代再提高5%~8%(Danell et al., 1993);欧洲赤松(Pinus sylvestris)和挪威云杉(Picea abies)每代种子园增益递增10%(Rosvall et al., 2004)。杉木3种世代种子园之间这种快速下降趋势,虽然是基于本试验的材料得出的,并有某些客观原因,但是也在一定程度上,反映出我国杉木种子园在向高世代过渡中,可能存在一些需要改进的潜在问题。我国的种子园,大多数具有育种群体与生产群体双重身份,其内在存在着保持遗传基础多样性与大幅提高遗传增益的矛盾(福克纳,1981),随着高世代种子园的发展,这种矛盾将制约着种子园遗传增益的增长,很有可能使遗传增益的世代间增幅进一步下降。纵观国际上先进的育种方案和建园经验,其中关键的一点,就是将育种群体与生产群体分开建立,现已取得良好效果,而且育种群体有了更深层次的发展(Cotterill, 1989;Danell, 1993;Powell et al., 2004; 马常耕,1993;1994;沈熙环,1994;王明庥,2001)。因此,我国在未来的种子园建设中,应该仔细研究和学习国际上先进的建园经验,制定长期育种策略,改变育种群体与生产群体合二为一的简单化方式,实行二者分别经营,从根本上改变这种状况。
陈代喜, 张文峤, 张晓东, 等. 1997. 杉木优良家系区域化试验研究. 广西林业科学, 26(1): 23-29. |
陈代喜, 黄开勇, 莫钊志, 等. 1999. 杉木改良代种子园遗传组成及效益评价. 广西林业科学, 28(2): 66-70. |
方乐金, 施季森, 张运斌, 等. 1998. 杉木优良家系及单株综合选择研究. 南京林业大学学报, 22(1): 17-20. |
福克纳R. 1981. 林木种子园. 徐燕千, 译. 北京: 中国林业出版社
|
国家标准总局. 1982. GB2772-81. 林木种子检验方法. 北京: 技术标准出版社
|
洪昌端, 陈天霞, 沈辛作, 等. 1992. 杉木优良杂交组合筛选及配合力分析. 浙江林业科技, 12(6): 1-8. |
李寿茂, 施季森, 陈孝丑, 等. 1999. 杉木第二代种子园自由授粉家系的评选. 南京林业大学学报, 23(4): 67-70. DOI:10.3969/j.issn.1000-2006.1999.04.015 |
李锦清, 董耀卿, 何秉云, 等. 1991. 浙江长乐杉木种子园营建技术和改良效果的研究. 林业科学研究, 4(1): 50-56. |
李锦清, 张建中, 汤志林, 等. 1990. 杉木种子园单亲本子代测定结果的分析. 浙江林学院学报, 7(1): 8-14. |
马常耕. 1993. 林木遗传改良发展中的新见解. 世界林业研究, (2): 13-22. |
马常耕. 1994. 高世代种子园营建研究的进展. 世界林业研究, (1): 31-38. |
马育华. 1982. 植物育种的数量遗传学基础. 南京: 江苏科学技术出版社.
|
全国杉木种源试验协作组. 1994. 杉木造林优良种源选择. 林业科学研究, 7(专刊): 1-25. |
阮梓材, 胡德活, 陈仲, 等. 1998. 杉木双亲本子代选择研究. 广东林业科技, 14(1): 1-9. |
沈熙环. 1994. 种子园优质高产技术. 北京: 中国林业出版社, 1-9.
|
孙鸿有, 郑勇平, 付顺华, 等. 2003. 杉木不同世代、类型种子园种子品质改良效果的研究. 南京林业大学学报, 27(2): 40-44. |
王欣. 1991. 杉木优良家系选择及1. 5代种子园建立技术的研究.贵州林业科技, 19(4): 9-19. |
王明庥. 2001. 林木遗传育种学. 北京: 中国林业出版社.
|
王赵民, 陈益泰. 1988. 杉木主要生长性状配合力分析及杂种优势的利用. 林业科学研究, 1(6): 614-624. |
吴隆高, 兰玉. 1993a. 11年生杉木全同胞子代林遗传效应研究. 浙江林学院学报, 10(4): 407-413. |
吴隆高, 张建章, 游顺昌, 等. 1993b. 十年生杉木初级种子园遗传效益分析. 浙江林业科技, 13(3): 22-25. |
徐金良, 洪昌端, 沈辛作, 等. 1997. 杉木不同类别良种改良效果的比较研究. 林业科学研究, 10(3): 337-340. DOI:10.3321/j.issn:1001-1498.1997.03.020 |
张建章, 兰玉, 祝春敏. 1991. 九年生杉木全同胞子代林的遗传分析. 浙江林业科技, 11(4): 23-29. |
张建忠, 徐永勤, 沈风强, 等. 2005. 杉木2代种子园单亲子代试验. 林业科学研究, 18(5): 632-635. DOI:10.3321/j.issn:1001-1498.2005.05.023 |
赵剑斌, 何祯祥. 1996. 杉木第二代种子园子代测定效果分析. 云南林业科技, 77(4): 41-45. |
周显昌, 张含国, 范玉清, 等. 1999. 加拿大林木遗传改良及其良种策略. 林业科技, 24(1): 7-9. DOI:10.3969/j.issn.1001-2117.1999.01.003 |
邹斌, 胡德活, 阮梓材, 等. 2003. 杉木第二代种子园效果分析. 华南农业大学学报, 24(4): 13-16. DOI:10.3969/j.issn.1001-411X.2003.04.004 |
Cotterill P P. 1989. The nucleus breeding system//Southern Forest Tree Improvement Committee. Proceedings of 20th southern forest tree improvement conference. Charleston, South Carolina, 36-42
|
Danell O, Ericsson T, Rosvall O. 1993. Currently suggested plans for long-term breeding of lodgepole pine and other conifers in Sweden//Lindgren D. Proceedings of "Pinus contorta-from untamed forest to domesticated crop" meeting of 1992 IUFRD WP 2. 02. 06. Umea, 271-278
|
Danell O. 1993. Breeding programmes in Sweden 1: general approach//Lee S J. Proceedings-Nordic group for tree breeding, progeny testing and breeding strategies. Edinburgh, Scotland, 6-10
|
Hodge G R, White T L, Powell G L, et al. 1989. Predicted genetic gains from one generation of Slash pine tree improvement. Sorth J Appl For, 13(1): 51-56. |
Lee S J. 1994. Breeding Radiata pine in New Zealand. Scottish Forestry, 48(1): 9-15. |
Matziris D I. 2000. Genetic variation and realized genetic gain from Aleppo pine tree improvement. Silvae Genetica, 49(1): 5-10. |
Matziris D I. 2005. Genetic variation and realized genetic gain from black pine improvement. Silvae Genetica, 54(3): 96-104. |
Powell M B. McRae T A, Wu H X, et al. 2004. Breeding strategy for Pinus radiata in Australia//Li Bailian, McKeand S. Proceedings of 2004 IUFRO joint conference of division 2: Forest genetics and tree breeding in the age of genomics, progress and future. Charleston, South Carolina, 308-318
|
Rosvall O, Stahl P H. 2004. New Swedish seed orchard program//Li Bailian, McKeand S. Proceedings of 2004 IUFRO joint conference of division 2: Forest genetics and tree breeding in the age of genomics, progress and future. Charleston, South Carolina, 141
|
Shelbourne C J A. 1991. Genetic gains from different kinds of breeding population and seed or plant production population. S Afr For, (160): 49-60. |
Squillace A E. 1989. Tree improvement accomplishment in the south//Southen Forest Tree Improvement Committee. Proccedings of 20th southern forest tree improvement conference. Charleston, South Carolina, 9-20
|
Steiner K C. 1993. Genetic improvement of oaks in North America. Ann Sci For, 50(Suppl 1): 359-367. |
Werner M. 1993. Breeding broadleaved tree species Sweden//Lee S J. Proceedings of Nordic group for tree breeding: Progeny testing and breeding strategies. Edinburgh, Scotland, 146-150
|
Wright J A, Osorio L F, Dvorak W S. 1996. Realised and predicted genetic gain in the Pinus patula breeding program of Smurfit cartor de Colombia. South African Forestry Journal, (175): 19-27. |