 2007, Vol. 43
2007, Vol. 43文章信息
- 红雨, 王林和.
- Hong Yu, Wang Linhe.
- 臭柏群落在不同演替阶段的PRO、MDA含量及保护酶活性变化
- Changes of PRO, MDA Contents and Protective Enzymes Activity of Sabina vulgaris Community at the Different Successional Stages
- 林业科学, 2007, 43(2): 115-119.
- Scientia Silvae Sinicae, 2007, 43(2): 115-119.
-
文章历史
- 收稿日期:2006-04-29
-
作者相关文章


2. 内蒙古农业大学生态与环境学院 呼和浩特 010022
2. Ecology and Environment College of Inner Mongolia Agricultural University Huhhot 010022
毛乌素沙地位于鄂尔多斯高原南部,海拔约1 300~1 600 m,年平均温度6~9 ℃,最热月7月平均温度20~24 ℃,最冷月1月平均温度-12 ℃~-8 ℃。年平均降水量250~490 mm,由东南向西北递减。毛乌素沙地是具有特殊地理景观的生态过渡带(Holt et al., 1997)。臭柏(Sabina vulgaris)为柏科圆柏属常绿匍匐针叶灌木,也是鄂尔多斯高原毛乌素沙地唯一的天然常绿灌木。多密集成片,根系发达,枝叶繁茂,耐风蚀沙埋,是优良珍稀的常绿固沙和护坡保土树种。在恶劣的沙地环境下, 仍然以高达70%~95%的覆盖度生存, 形成占绝对优势的单优群落(张国盛等,2001)。国内外的研究者针对其生物生态学特性(松尾奈绪子等, 2001)、生育规律(王林和等,1998;2002;何维明,2002)、繁殖(山中典和等,2000)、遗传多样性(红雨等,2006a;2006b)等特性进行了大量的研究。但对臭柏群落演替及不同演替阶段的生理反应和适应性反应研究甚少。笔者对不同演替阶段臭柏群落生理反应和适应性反应进行了研究, 探讨毛乌素沙地天然臭柏群落在不同演替阶段对沙地环境的生态适应性。
1 材料与方法 1.1 材料的采集2004年6—8月,在毛乌素沙地图克乡北试验地上采用空间代替时间的方法选取了臭柏演替各阶段(朱志诚等,1996;郭柯,2000)的典型样地。样品采集群落及采样数目见表 1。不同演替阶段,随机采集臭柏新鲜叶片,叶位、方向及采集时间基本一致,一部分叶片在65 ℃烘干24 h后测定脯氨酸(PRO)含量,一部分用液氮固定,回实验室后转入-80 ℃的冰箱用于丙二醛(MDA)含量和超氧化物歧化酶(SOD)、过氧化物酶(POD)和过氧化氢酶(CAT)活性的测定。
|
|

1) PRO的测定 测定方法参见张志良等(2003)。取已经称重的臭柏叶片0.5 g放入已经用清水洗净并擦拭干净的研钵中,加入8 mL 3%的磺基水杨酸研磨,研磨后的溶液倒入离心管中,塞上塞子静置,提取上清液。把提取的上清液放入沸水浴锅中水浴10 min后冷却。把冷却后的离心管内上清液在天平上平衡后,在3 000 r·min-1下离心10 min,取上清液待测。用移液管吸取上清液2 mL,2 mL冰醋酸,4 mL 2.5%茚三酮溶液于试管中,对照为2 mL蒸馏水,2 mL冰醋酸,4 mL 2.5%的茚三酮溶液,沸水浴60 min后冷却。用移液管加4 mL甲苯于冷却后的上清液中,振荡后静置分层。取上层PRO红色甲苯溶液于比色杯中,在723型分光光度计520 nm波长处比色。记取测得的吸光度值。
2) MDA、保护酶的测定 a)提取液的制备 取冷藏于-80 ℃的臭柏叶片1 g,加入磷酸缓冲液(PBS,pH7.8)研磨至匀浆。4 ℃,10 000 r·min-1离心15 min, 取上清液备用。b)MDA的测定 测定方法参见张志良等(2003)。吸取1 mL提取液,加入1 mL PBS(62.5 mmol·L-1, pH7.8),再加入2 mL 10%三氯乙酸(TCA)[含0.5%硫巴比妥酸(TBA)],混匀,在试管上加盖塞,置于沸水浴中沸煮15 min,迅速冷却,4 000 r·min-1离心10 min,以10%TCA(含0.5%TBA)为对照,取上清液测定450 nm、532 nm和600 nm下的OD值。MDA浓度=6.45(D532-D600)-0.56D450。c) SOD活性的测定 测定方法参见张志良等(2003)。反应介质(50 mmol·L-1 pH7.8 PBS,内含77.12 μmol·L-1 NBT,0.5 mmol·L-1 EDTA, 13.37 mmol·L-1蛋氨酸)3.9 mL,待测酶液10 μL,80.2 μmol·L-1核黄素0.1 mL,以不加待测酶液(以缓冲液代替)的试管为对照,然后将各管在400 lx下反应10 min进行光化还原。反应酶在560 nm处测定OD值,以抑制光化还原50%作为一个酶活性单位(U),酶活性以U·g-1DW表示。d) POD活性的测定 测定方法参见张志良等(2003)。反应系统:2.0 mL愈创木酚溶液(250 mL 0.1 mol·L-1 pH6.0 PBS+0.74 mL愈创木酚配制),50 μL待测酶液和20 μL 30% H2O2。加入1 min H2O2后比色波长470 nm,室温。以愈创木酚溶液做对照。在470 nm处OD值每分钟增加0.01为一个酶活性单位(U),酶活性以U·g-1DW min-1表示。e) CAT活性的测定 测定方法参见张志良等(2003)。5 mL反应液中含有PBS 50 mmol·L-1 (pH7.0),H2O2 8 mmol·L-1及100μL待测酶液。室温下反应1 min,加2 mL 10% H2SO4终止反应。用2 mmol·L-1 KMnO4滴定剩余的H2O2,根据H2O2的消失量计算CAT活性。酶活性单位为μmol H2O2·g-1DW min-1。
2 结果与分析 2.1 不同演替阶段PRO含量的变化据图 1显示,演替过程中臭柏PRO含量呈先上升后下降再上升的变化。方差分析显示,臭柏的PRO含量在阶段间差异极显著。进一步多重比较显示,演替的第Ⅲ阶段与演替的第Ⅰ阶段、演替的第Ⅱ阶段、第Ⅳ阶段差异达到了极显著水平。在半固定沙地油蒿+臭柏群落(Ⅰ)向固定沙地臭柏+硬质早熟禾群落(Ⅱ)发展时期, 沙地也由半固定沙地演变为固定沙地,相应的含水量增加,肥力升高。随着环境的改善,植物体内不应该累积大量的PRO,但是试验数据表明在此阶段臭柏PRO含量迅速升高。笔者在采集材料的过程中发现此阶段的臭柏枝条上有较多球果,据Tang(1989)的研究表明PRO的积累可稳定大分子化合物,对植物组织分化和发育有重要作用。在此阶段PRO的积累可以维持叶渗透压和保水力,另外对蛋白质的稳定性也有显著影响,从而使臭柏在繁殖时期没有受到生理伤害,这对臭柏的生存和生长起到了重要作用,可能是一种适应性反应。但是,是否与上述原因有直接关系还需进一步研究。

|
图 1 不同演替阶段臭柏PRO、MDA含量的变化 Fig. 1 Changes of the PRO, MDA contents under different successional stages\= 字母不同者表示差异极显著(P<0.01),“-"表示没有差异。Different letters in the same picture indicates very significant difference. “-" indicates no significant difference.下同。The same below. |
随着演替的进行,臭柏+硬质早熟禾群落进展到臭柏群落,此时臭柏群落处于演替亚顶极时期(红雨,2006b),这一时期表现为群落结构的复杂化,地上和地下空间的最大利用,生产力的最大利用和生产率的增加,群落生境的中生化和群落环境的强烈改造,此时地表形成一层4~7 cm厚的沙质壤土,一般不超出10 cm,基质物理和化学特征以及水条件向着有利于植物生活的方向发展,因此PRO积累较少。臭柏群落继续发展为臭柏+苔藓群落,这一时期地表形成结皮,雨水很难透入土壤,大量被流失。PRO作为一种良好的渗透调节物质,它可使臭柏在水分亏缺时降低其渗透势,从外界水势低的环境中继续吸水,同时还有助于细胞核组织的持水,防止脱水,以增强臭柏的耐旱性,所以可以说臭柏体内PRO含量的大量累积与其抗旱性之间存在着密切的关系。
2.2 不同演替阶段MDA含量的变化不同演替阶段的臭柏叶片中MDA的含量呈现微小的上升、下降和较显著的再上升趋势。方差分析显示,臭柏MDA含量阶段间差异极显著。进一步多重比较显示,演替第Ⅰ阶段、第Ⅱ阶段和演替第Ⅲ阶段的MDA含量变化不大,阶段间差异不显著,但演替的前3个阶段均与演替第Ⅳ阶段呈现极显著的差异水平(图 1)。在演替第Ⅰ阶段,半流动沙地上土壤含水量较少,土壤紧实度较低,而且土壤较贫瘠,所以演替第Ⅰ阶段MDA的含量要比第Ⅲ阶段略高一些。在演替第Ⅳ阶段MDA含量迅速增长,表明细胞膜脂过氧化程度增加,脂膜受伤害程度也在加深。
2.3 不同演替阶段SOD活性的变化根据图 2显示,臭柏随着演替阶段的不同SOD活性呈现微小的上升、下降和较显著的再上升趋势。在演替的第Ⅱ阶段有所上升,在演替Ⅳ阶段活性增高,与其脯氨酸和丙二醛含量的变化趋势相吻合,是一种环境胁迫下的一种保护性适应。方差分析显示,臭柏SOD含量阶段间差异极显著。进一步多重比较显示,臭柏演替第Ⅱ阶段与第Ⅲ阶段呈现显著差异,说明生理胁迫后SOD有较明显的升高。演替第Ⅳ阶段,酶活性的升高较明显与前3个阶段呈现极显著的差异水平,可以推测演替第Ⅳ阶段对于臭柏的生存、生长起到了质的关键性作用。许多研究表明,SOD活性于植物的抗旱性密切相关。在水分胁迫下抗旱性强的品种或种具有较高的SOD活性,这已在沙米(Agriophyllum pungens)和欧亚旋覆花(Inula britanica)、差不嘎蒿(Artemisia halodendron)、白草(Pennisetum centrasiaticum)、狗尾草(Setaria viridis)(周瑞莲等,1999a; 1999b)、牛心朴子(Cynanchum komarovii)(米海莉等,2003),对干旱胁迫下几种荒漠植物沙拐枣(Calligonum mongolicum)、梭梭(Haloxylon ammodendron)和沙枣(Elaeagnus angustifolia)(龚吉蕊等,2004),小麦(武宝轩等, 1985),水稻(吴岳轩等,1994)等植物上得到证实。沙漠化环境中的植物趋向于抗干旱。本试验研究表明,臭柏的抗干旱能力很强,其SOD活性增幅也较大,属于依赖保护酶进行抗逆的植物类型。

|
图 2 不同演替阶段SOD、POD、CAT活性变化 Fig. 2 Changes of the SOD、POD、CAT activity under different successional stages |
根据图 2可知,在演替过程中,臭柏的POD活性呈先下降再上升趋势,在演替第Ⅲ阶段出现最低值,在演替第Ⅳ阶段又迅速上升。与其PRO、MDA含量和SOD活性等的变化相吻合。表明在演替第Ⅲ阶段水分、养分等条件较好,环境胁迫有所缓解,所以POD活性较稳定,没有出现升高现象。在演替第Ⅳ阶段,一方面可能是由于胁迫的加重,体内H2O2的含量高诱导了POD活性的增高,另一方面也许叶片中的脯氨酸的大量积累激发了POD活性的增加。说明在干旱胁迫下SOD、POD协同作用在沙生植物抗膜脂过氧化,保护膜的完整性上起重要作用。方差分析显示,演替阶段间差异极显著。通过阶段间的多重比较发现,臭柏演替第Ⅳ阶段分别与演替前3个阶段间差异极显著,其余阶段间差异不显著。
2.5 不同演替阶段CAT活性的变化如图 2所示,不同演替阶段CAT活性变化不大,略有上升的趋势。方差分析显示,叶片中的POD活性在不同演替阶段间差异不显著。随着演替的发展,演替后期SOD、POD等都有升高的现象,而CAT没有升高保持了原有水平。可能是在环境胁迫下原来积存的CAT等就已经能够清除过氧化物体和线粒体中的反应性氧(ROS),所以在演替第Ⅳ阶段,环境胁迫下没有诱导CAT,使CAT活性保持了原有水平。
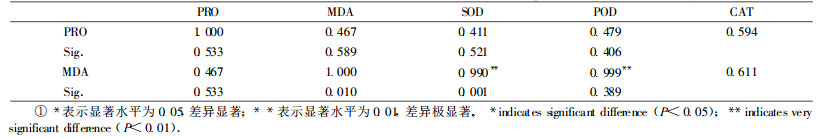
2.6 PRO含量、MDA含量与保护酶活性的相关性分析根据表 2显示,SOD活性和POD活性变化与MDA含量的变化呈极显著的正相关。说明MDA含量增加,SOD、POD活性迅速增加。
|
|
臭柏群落的演替过程中,PRO、MDA含量呈先上升后下降再上升的变化,各演替阶段间差异极显著。SOD、POD活性在各演替阶段间差异极显著,而CAT活性差异不显著。不同植物3种保护酶互相配合的形式不同。在演替第Ⅲ阶段环境胁迫相对较小,臭柏体内的ROS相对较少,膜脂过氧化程度较低,SOD活性较为稳定,没有增高;在演替第Ⅳ阶段环境胁迫增大,臭柏体内的ROS增加,膜脂过氧化作用加强,MDA含量增加,SOD、POD活性迅速增加。POD可能是旱生植物臭柏茎、叶清除H2O2的主要酶。它与SOD协同作用在抑制膜脂过氧化作用上起重要作用。在演替第Ⅳ阶段只有POD活性的增幅较大,CAT活性没有显著变化,导致了在演替第Ⅳ阶段衰老细胞通过各种代谢途径生成的H2O2比降解掉的H2O2多,所以在演替第Ⅳ阶段H2O2含量增多可能是MDA含量增高的原因。
演替第Ⅳ阶段游离PRO含量的上升,MDA含量的增加,SOD、POD活性增加,表现出环境胁迫下植物受损的生理共性。从各项指标来看,一是多数指标在演替第Ⅲ阶段出现较低水平,因此可以看出在演替第Ⅲ阶段是环境最稳定的关键时期;二是在演替第Ⅳ阶段臭柏体内出现一系列的受损表现。伴随着演替过程的进行,环境的变化过程中有机体的生理作用并非是单一的,而是酶协调一致共同清除活性氧的侵害。总之,臭柏在对恶劣环境的长期适应过程中产生了一系列的生理适应机制。
龚吉蕊, 赵爱芬, 张立新, 等. 2004. 干旱胁迫下几种荒漠植物抗氧化能力的比较研究. 西北植物学报, 24(9): 1570-1577. DOI:10.3321/j.issn:1000-4025.2004.09.002 |
郭柯. 2000. 毛乌素沙地油蒿群落的循环演替. 植物生态学报, 24(2): 243-247. DOI:10.3321/j.issn:1005-264X.2000.02.020 |
何维明, 张新时. 2002. 沙地柏对毛乌素沙地3种生境中养分资源的反应. 林业科学, 38(5): 1-6. DOI:10.3321/j.issn:1001-7488.2002.05.001 |
红雨, 王林和, 梁小荣. 2006a. 不同生境臭柏种群的遗传多样性分析及其与环境因子的相关性. 干旱区资源与环境, 20(3): 184-187. |
红雨, 王林和, 张国盛, 等. 2006b. 臭柏种群在不同演替阶段的遗传多样性分析. 应用生态学报, 17(11): 2006-2010. |
米海莉, 许兴, 李树华, 等. 2003. 干旱胁迫下牛心朴子幼苗相对含水量、质膜透性及保护酶活性变化. 西北植物学报, 23(11): 1871-1876. DOI:10.3321/j.issn:1000-4025.2003.11.003 |
王林和, 张国盛, 董智. 1998. 毛乌素沙地臭柏种子产量及更新的初步研究. 林业科学, 34(6): 105-112. DOI:10.3321/j.issn:1001-7488.1998.06.014 |
王林和, 刘美珍, 张国盛, 等. 2002. 毛乌素沙地臭柏不定根发生特性的研究. 林业科学, 38(5): 156-159. DOI:10.3321/j.issn:1001-7488.2002.05.026 |
武宝轩, 格林·托得. 1985. 小麦幼苗中超氧化物歧化酶活性与幼苗脱水忍耐力相关性的研究. 植物学报, 27(2): 152-160. |
吴岳轩, 吴振球. 1994. 水稻幼苗超氧化物歧化酶器官特异性的研究. 湘潭师范学院学报, 15(3): 43-47. |
张国盛, 高润宏, 王林和, 等. 2001. 毛乌素沙地臭柏群落结构和生物多样性组成研究. 内蒙古农业大学学报, 22(4): 88-91. |
张志良, 瞿伟菁. 2003. 植物生理学实验指导. 北京: 高等教育出版社, 121-258.
|
周瑞莲, 王海欧. 1999a. 在干旱、高温胁迫中沙生植物抗脱水性与膜脂过氧化关系的研究. 中国沙漠, 19(增1): 59-64. |
周瑞莲, 赵哈林, 王海鸥. 1999b. 科尔沁沙地植被演替的抗逆性特征. 中国沙漠, 19(1): 1-6. |
朱志诚, 郭爱莲, 岳明. 1996. 陕北黄土高原臭柏群落进展演替和逆行演替. 西北大学学报:自然科学版, 26(4): 325-330. |
山中典和, 王林和, 吉川贤. 2000.  .日本绿化工学会志, 25(4): 437-442 .日本绿化工学会志, 25(4): 437-442
|
松尾奈绪子, 大手信人, 木庭啟介, 等. 2001.  析.日本绿化工学会志, 27(1): 68-76 析.日本绿化工学会志, 27(1): 68-76
|
Holt J A. 1997. Grazing pressure and soil carbon, microbial biomass and enzyme activities in semiarid Northeastern Australia. Applied Soil Ecology, 5: 143-149. DOI:10.1016/S0929-1393(96)00145-X |
Tang Zhangcheng. 1989. The accumulation of free proline and its roles in water-stressed sorghum seedings. Acta Phytophsiological Sinica, 15(1): 105-110. |