 2007, Vol. 43
2007, Vol. 43文章信息
- 李涵, 殷云龙, 徐朗莱, 於朝广, 陈永辉.
- Li Han, Yin Yunlong, Xu Langlai, Yu Chaoguang, Chen Yonghui.
- 落羽杉属树种及其杂交后代亲缘关系的RAPD分析
- Genetic Relationships of Taxodium Species and Their Hybrids Basedon RAPD Analysis
- 林业科学, 2007, 43(2): 48-51.
- Scientia Silvae Sinicae, 2007, 43(2): 48-51.
-
文章历史
- 收稿日期:2005-12-07
-
作者相关文章



2. 南京农业大学生命科学学院 南京 210095
2. College of Life Sciences, Nanjing Agriculture University Nanjing 210095
落羽杉属(Taxodium)为杉科(Taxodiaceae)落叶或半常绿性乔木,本属有3个种,分别是落羽杉(T. distichum)、池杉(T. ascendens)和墨西哥落羽杉(可简称为墨杉)(T. mucronatum),具生长快、干型直、材质好和适应性广的特点,是林业和园林建设中很有发展前景的树种。江苏省·中国科学院植物研究所从1973年起,先后开展了落羽杉属种间杂交试验和速生耐碱类型的选育(陈永辉等,1988; 1989),获得速生耐碱无性系中山杉302(T. distichum×T. mucronatum),于2002年通过国家林业局林木品种审定委员会审定,成为全国首批通过国家审定的16个林木良种之一。20世纪90年代又进行了以中山杉302为母本、墨杉为父本的回交选育(殷云龙等,2003),选出的中山杉回交一代(BCF1)102、118、149于2004年通过江苏省林业局林木良种审定委员会的认定。
随着人们对中山杉利用价值的不断认识,其推广应用规模迅速扩大(殷云龙等,2005),中山杉302及其回交代均表现出明显的杂种优势,而且回交代各无性系在性状表现上变异很大(陈永辉等,2006),从而说明了研究落羽杉属树种及其杂交后代的遗传变异及亲缘关系的必要性。周康(1998)从形态上分析了落羽杉属树木及其F1代杂种的遗传和变异;尹晓明等(2002)以中山杉302和墨杉及其回交一代中BCF161、102、118、149为材料,用同工酶电泳方法分析回交杂种之间同工酶酶谱的异同。但植物形态标记受环境因素影响较大,同工酶标记也受基因表达的限制而影响其使用。分子标记技术的发展为研究植物亲缘关系提供了有效途径,其中RAPD分子标记技术自1990年(Williams et al., 1990)建立以来已得到了广泛的应用。由于它具有快速、简便的特点,在严格的试验条件下有良好的重复性,并且1次试验提供较多的分析位点,因此可以作为一种分子遗传标记来识别物种之间的亲缘关系,目前该技术已在林木遗传育种研究中广泛应用。李春香等(1999)利用RAPD技术研究了水杉(Metasequoia glyptostroboides)27个样本的遗传多样性;陈云鹏等(2002)利用RAPD检测技术推测杂交墨杉(墨杉×柳杉)(T. mucronatum×Cryptomeria fortunei)的亲缘关系;邓白罗等(2006)利用RAPD技术研究了山茶属(Camellia)红山茶组(Section Camellia)29种植物的亲缘关系及分类。本研究利用RAPD技术对落羽杉属3个种及其杂交后代进行遗传多样性的分析,旨在进一步明确它们之间的亲缘关系,为今后育种提供可靠依据。
1 材料和方法 1.1 材料南京中山植物园内落羽杉和墨杉均为10年生嫁接苗,池杉为27年生嫁接苗;中山杉302(落羽杉×墨杉)(F1)为10年生嫁接苗,回交后代(中山杉302×墨杉)BCF11、BCF127、BCF186、BCF1102、BCF1118、BCF1136、BCF1140、BCF1149、杂交墨杉(墨杉×柳杉)均为1年生嫁接苗,种植于南京中山植物园种质资源圃。分别取营养枝上的新鲜幼嫩叶片,每种材料随机选取多个个体(同一无性系扦插苗)进行采样,冰盒保鲜带回实验室,用双蒸去离子水冲洗干净,放入塑料袋中置于-80 ℃冰箱备用。
1.2 基因DNA提取按照陈云鹏等(2002)的方法稍加改进提取DNA。取200 mg叶片,放入研钵中,迅速加入液氮充分研磨成粉,装在1.5 mL离心管中;加入1 mL DNA提取缓冲液Ⅰ(50 mmo1·L-1 Tris-HCl,5 mmo1·L-1 EDTA,350 mmo1·L-1山梨醇,0.1%β-巯基乙醇,10%PEG6000,pH8.0),充分混匀后,以12 000 r·min-1于4 ℃下离心5 min;倒去上清液,沉淀重悬于0.5 mL预热的DNA提取缓冲液Ⅱ(1%月桂酰基肌氨酸钠,710 mmo1·L-1 NaCl,0.1%CTAB,其余成分同DNA提取缓冲液Ⅰ);放入60 ℃水浴中25~30 min,加入等体积的氯仿:异戊醇(体积比为24:1),小心混匀15 min,室温下以12 000 r·min-1离心15 min;上清液转入新的1.5 mL离心管,加入50 μL 10%CTAB/0.7 mo1·L-1 NaCl溶液,置65 ℃的水浴中10 min,加入10 μL Rnase 37 ℃处理30 min,再加入等体积的氯仿:异戊醇(体积比为24:1),小心混匀10 min,室温下以12 000 r·min-1离心15 min(若上清液粘稠浑浊,重复10%CTAB/0.7 mo1·L-1 NaCl抽提步骤);取上清液,加入0.7倍体积预冷的异丙醇(-20 ℃),轻轻摇匀,将离心管置于冰箱中沉淀DNA 5~10 min;以12 000 r·min-1于4 ℃下离心5 min,DNA沉淀用800 μL 70%乙醇洗涤2次,自然风干后,溶于40 μL TE中。
1.3 AP-PCR反应体系组成及扩增程序选用10碱基寡聚核苷酸随机引物,反应体积20 μL:10×PCR缓冲液(终浓度1×),2.5 mmo1·L-1 MgCl2,0.25mmo1·L-1 dNTPs,24 ng引物(Invitrogen公司合成),10~20 ng模板,1.2 U Taq DNA聚合酶(Takara公司)。反应程序为:于94 ℃中预变性4 min;再在94 ℃中变性0.5 min,35 ℃中复性1 min,72 ℃中延伸1.5 min,45个循环;最后一步72 ℃延伸7 min。
PCR产物在1.5%琼脂糖凝胶上电泳,电泳缓冲液为1×TAE,在凝胶中加入0.5 μg·mL-1 EB,电泳完毕后在紫外灯下观察照相。
1.4 数据处理与分析选择重现性好且稳定的扩增反应谱带进行系统分析,每条谱带作为1个性状,重现性强或亮度强的记为1,重现性弱、亮度差或无带的记为0。根据公式S=2Nxy/(Nx+Ny)×100%(式中:Nxy表示X种和Y种共同具有的RAPD条带,Nx与Ny分别为X种和Y种所具有的RAPD条带)计算出任意2个个体间的相似系数,遗传距离D=1-S。利用SPSS11.0程序进行聚类分析。
2 结果与分析 2.1 引物筛选及扩增结果对100个引物进行筛选,从中选出扩增条带清晰的13个引物对所有的样本进行扩增,每个引物至少重复扩增1次以肯定重复性。共扩增出DNA带92条,其中72条具多态性,占总带数的78.26%,平均每个引物扩增的DNA带数为7.1条,多态性DNA的带数为5.5条(图 1)。

|
图 1 引物S1003(上)、引物S1014(下)扩增的RAPD图谱 Fig. 1 RAPD bands amplified with primer S1003 (upper) and S1014 (below) 1.落羽杉T. distichum; 2.墨杉T. mucronatum; 3.池杉T. ascendens; 4.F1302 (T. distichum×T. mucronatum); 5.BCF11; 6.BCF127;7.BCF186; 8.BCF1102; 9.BCF1118; 10.BCF1136; 11.BCF1140;12.BCF1149; 13.杂交墨杉T. mucronatum×Cryptomeria fortunei. |
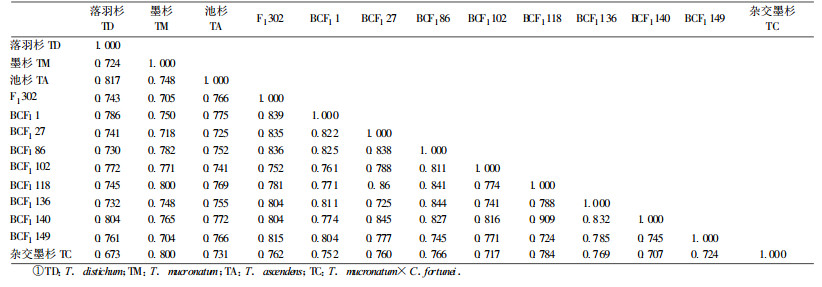
13个引物的扩增产物经聚类分析,获得不同样本的遗传相似系数矩阵(表 1),基于RAPD结果进行聚类分析所得的树状图见图 2。通过比较可以发现,3个原种之间,落羽杉和池杉的相似性较高(S=0.817),墨杉和它们之间相似系数较低,说明落羽杉和池杉的亲缘关系较近,与墨杉的亲缘关系稍远,这与殷云龙等(2003)的研究结果一致。中山杉302与母本落羽杉的相似系数为0.743,与父本墨杉相似系数为0.705,说明中山杉302更接近于母本,其形态上的差别也反映了这一点。在回交后代中,BCF1118与BCF1140相似系数最大;BCF11与母本F1302亲缘关系最近;BCF1118与父本墨杉亲缘关系最近。从聚类图可以看出,在阈值为5.2时,回交一代可以聚为3类:BCF1149和BCF1102各为一类,BCF11、27、118、140、86、136聚为一类。
|
|

|
图 2 根据RAPD结果构建的供试样本的聚类图 Fig. 2 The clustering dendrogram for the samples based on RAPD data |
在对中山杉302回交一代的早期选育研究中,BCF11、118、140这3个无性系,总体性状与中山杉302相似,干型直,枝叶密,秋叶变色期较迟,落叶期晚,半常绿性状在苗期表现较突出;BCF1102无性系总体性状与羽叶池杉(T. ascendens cv. ‘Yuyechisha')相似,顶端生长优势明显,而粗生长较池杉快,针叶相对较短,在脱落性小枝上着生角度较小(<45°);BCF1149无性系总体性状与落羽杉相似,顶端优势不强,但枝叶浓密,粗生长较快,秋末冬初其叶色呈棕红色,落叶期较落羽杉迟。本试验的聚类结果与杂交后代的形态和性状表型分类基本一致。从聚类图可以看出杂交墨杉(墨杉×柳杉)与墨杉亲缘关系很近,两者的相似系数为0.800,由于未将杂交墨杉的父本柳杉进行RAPD分析,故不能断定杂交墨杉与哪一个亲本的亲缘关系较近,这与陈云鹏等(2002)研究结果一致。
连续3年对回交一代无性系的苗期生长量(株高和地径)、标准木的地上部分生物量和分枝结构的测定结果(殷云龙等,2003)表明,BCF1118的生长量和地上部分生物量明显高于亲本中山杉302,其树干生物量占地上部分总生物量的比例大于50%,而树枝生物量比例较低,对盐碱地具有很好的适应性,可以作为速生用材类型;BCF1149由于叶色红,与亲本叶片绿色不同,且在冬季树叶仍呈鲜红色,可以作为速生观赏类型。在回交后代中,BCF1118与父本墨杉的亲缘关系最为接近,可以作为新的杂交选配应用于以后的育种工作中。由聚类图可以看到,在回交后代中BCF1149与双亲之间相距较远,综合其外部形态和生理特性,说明BCF1149在继承了双亲遗传物质的同时还表现出一定程度的变异,在以后的杂交育种工作中也具有潜在的利用价值。
Nei(1978)认为,对所用样本数较少的群体,通过分析大量的基因位点可以抵消由样本数较少引起的误差。而增加引物的数目是RAPD获得大量基因位点的重要措施。本试验中所筛选出的引物数目相对来说还不够,为了获得更有价值、更为精确的结果,第一要扩大引物筛选的范围(如200条),从更多的随机引物中选出更多更好(如20条以上)能进行扩增的引物,这样在进行结果分析时更有说服力。但是RAPD所能提供的是显性标记,无法区分纯合型和杂合型基因,部分遗传信息难以表达。因此如果仅用RAPD一种标记进行研究,会在遗传分析上受到很多限制,并且获得的遗传信息也不全面。要解决这个问题,就需要其他分子标记技术如ISSR、RFLP、AFLP的配合。
陈永辉, 王名金, 伍寿彭. 1988. 落羽杉属的引种和选育. 江苏林业科技, 15(2): 43-49. |
陈永辉, 王名金, 伍寿彭, 等. 1989. 中山杉302和401无性系在碱地上的生长和适应性的初步研究. 江苏林业科技, 16(3): 14-18. |
陈永辉, 伍寿彭, 李永荣, 等. 2006. 落羽杉中山杉系列新品种选育初报. 江苏林业科技, 33(4): 1-5. DOI:10.3969/j.issn.1001-7380.2006.04.001 |
陈云鹏, 潘士华, 张建军, 等. 2002. 利用RAPD检测技术推测杂交落羽杉群落间的亲缘关系. 复旦学报:自然科学版, 41(6): 641-645. |
邓白罗, 谭晓风, 漆龙霖, 等. 2006. 山茶属红山茶组植物的RAPD分析及分类研究. 林业科学, 42(5): 36-41. |
李春香, 杨群, 周建平, 等. 1999. 水杉自然居群遗传多样性的RAPD研究. 中山大学学报:自然科学版, 38(1): 59-63. |
尹晓明, 殷云龙, 陈永辉. 2002. 中山杉302和墨西哥落羽杉及其回交一代的同工酶分析. 植物资源与环境学报, 11(3): 59-61. DOI:10.3969/j.issn.1674-7895.2002.03.013 |
殷云龙, 尹晓明, 於朝广, 等. 2003. 中山杉302回交一代的早期选育. 植物资源与环境学报, 12(2): 22-27. DOI:10.3969/j.issn.1674-7895.2003.02.006 |
殷云龙, 於朝广. 2005. 中山杉——落羽杉属树木杂交选育. 北京: 中国林业出版社, 140-144.
|
周康.1998.落羽杉属种间杂种的遗传分析及耐碱生理.中国科学院植物研究所硕士学位论文
|
Nei M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89: 583-590. |
Williams J G K, Kubelik A R, Livak K J, et al. 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res, 18(22): 6531-6535. DOI:10.1093/nar/18.22.6531 |