 2007, Vol. 43
2007, Vol. 43文章信息
- 康文星, 邓湘雯, 赵仲辉.
- Kang Wenxing, Deng Xiangwen, Zhao Zhonghui.
- 林冠截留在杉木林生态系统能量转换过程中的作用
- Effects of Canopy Interception on Energy Conversion Processes in Chinese Fir Plantation Ecosystem
- 林业科学, 2007, 43(2): 15-20.
- Scientia Silvae Sinicae, 2007, 43(2): 15-20.
-
文章历史
- 收稿日期:2006-01-04
-
作者相关文章


林冠是森林群落结构的重要特征之一,也是森林生态系统与周围环境进行能量和物质交换的重要场所。林冠的截留作用是森林生态系统一个重要的功能过程。许多学者对林冠的截留作用进行了多方面研究,如光在林冠内的传播(刘家冈等,1985),林冠层的光吸收和光分布规律(朱劲伟等,1982),林冠对大气降水的截留作用(谢春华等,2002;卫正新等,1997;文仕知等,1989;周光益等,1995;陈引珍等,2005)。不少学者也开展了林冠对降雨动能影响的研究(雷瑞德,1994;周国逸,1997;王彦辉,1986;中野秀章,1985)。然而,对于林冠截留在系统能量转换过程的功能作用,却少有报导。本文以连续定位的测定数据,分析了辐射能在林冠层的分配格局,林冠截留水分蒸发耗能规律和林冠对降雨动能的影响。以揭示林冠截留对森林生态系统能量转化和平衡的影响。
1 研究地概况本研究在国家重点野外科学观测站——中南林学院会同生态站第二集水区内进行的。生态站位于湖南会同县广坪深冲林场(26°50′ N,109°45′ E),年均气温16.7 ℃,年降水量1 100~1 600 mm,相对湿度80%以上,典型亚热带湿润气候。海拔270~400 m,地貌为低山丘陵,土壤为山地黄壤。研究的林分为杉木(Cunninghamia lanceolata)人工林纯林,密度为2 000株·hm-2,郁闭度为0.9,林分平均高度16 m,平均胸径15.5 cm,枝下高平均7 m。
2 研究方法将DFY2型天空辐射表、DFY3型直接辐射表安装在林冠表面上和林冠下,测定到达林冠的总辐射量、反射量以及穿过林冠的辐射量。林冠吸收辐射量用总辐射量减去反射量,再减去穿过林冠的辐射量求得。
在观测铁架上,用SL-1型遥测雨量计测定冠上大气降水。林冠下设2个18~20 m2的穿透水承接装置。用SW40型日记水位仪自动测定穿过林冠层的水量。采用聚乙烯塑料管蛇形缠绕树干基部,其下端插入一个容器内,测定树干流。林冠截留通过测定的大气降水减去测定的穿透水和树干流求得。林冠截留水分蒸发耗能用林冠截留水量乘上每克水变成相同温度的水汽所需热量(潜热)求得。
在林外和林内都设1个薄薄的石灰层和1个过了细筛的风干土层,面积都为1 m2。下雨时,即刻观测雨滴直径和分布。
3 结果与分析 3.1 林冠截留在辐射能分配中的作用表 1表明,林冠反射和吸收量的月变化规律与总辐射量的月变化规律一致,即总辐射量大的月份,林冠的吸收和反射量也大,总辐射量小的月份,林冠的吸收和反射量也小。1年中反射率出现2个低峰期:4—5月(反射率只有7%)和9—10月份(反射率只有7%~8%)。高峰期出现在气温高的月份,尤其是8月,高达11.3%。反射率的2个低峰期(4—5月和9—10月)与杉木生长的高峰期几乎同步,生长季节杉木生长的低峰期(8月)与反射率高峰值完全重合。可见,冠层反射率的变化与杉木生长发育密切相关。
|
|

从表 1中还可以看出,冠层对辐射的吸收率和透射率取决于林冠层的结构特征和枝叶的发育状况。冬半年,林木休眠,叶子变老,叶质变硬,颜色变深暗,对辐射的截留和吸收能力增强,林冠的透射率小(6.2%~6.3%)。生长季节,老叶脱落,新叶没有完全展开,从而减少了阻拦和吸收辐射的作用面(叶面积),林冠的透射率增大。这种现象在新老叶交换期的4—5月最为突出,透射率为9.6%~10.5%。
每年到达杉木林林冠的总辐射量为30.816 8亿J·m-2,林冠全年吸收的辐射量为25.543 0亿J·m-2,占投入辐射总量的82.9%,透过林冠层到达地面的辐射量只有2.530 6亿J·m-2,约为总辐射量的8.2%。林冠层的反射量2.743 2亿J·m-2,为总辐射的8.9%。杉木林冠层以它巨大的作用面(叶面积指数8)和重叠的枝叶(冠层厚度平均9 m)阻拦和吸收辐射能流,极大地减少了到达林冠下空间的辐射量,从而改变了杉木人工林生态系统内部能量,造就了林下空间的小气候环境。
3.2 林冠截留在系统水分蒸发耗能中的作用林冠截留水分的蒸发,有一个液态水变为气态水的过程,这种相变过程需要消耗能量。集水区杉木林冠每年截留水量261.60 mm,这些水分蒸发消耗能量6.369 5亿J·m-2,占杉木林生态系统获得的净辐射能(投入到林冠层表面的辐射能减去林冠层反射的能量)的22.9%。
用水量平衡法求出杉木林生态系统每年总蒸发散的水量862.03 mm,由此而消耗的能量20.952 4亿J·m-2。林冠截留水分蒸发消耗的能量为6.369 5亿J·m-2,占系统总蒸发散耗能的30.4%。在杉木林生态系统潜热能的支出中有近1/3是林冠截留水分蒸发耗能贡献的。
从表 2可看出,温度低、降水频繁且强度不大的2—4月,林冠截留水分蒸发耗能分别占该月净辐射能的38.8%~47.8%,占该月总蒸发散耗能的49.6%~55.6%,是该时期森林生态系统能量支出的最大项目。可见,越是温度低、多雨(小雨为主)的季节,越能显示出林冠截留在森林生态系统能量平衡中的重要作用。
|
|

雨滴动能是裸露土地发生侵蚀的主要动力,一些研究者根据自己的试验得出了一些计算雨滴动能的经验公式(Salles et al., 2002;Kinnell,1981;Ulbrich et al., 1985;徐锐,1983;蔡丽君,2003a; 2003b;贡力,2005;周佩华等,1981;王万忠,1987;江忠善等,1983;Disrud,1970),并普遍认为林冠能削弱雨滴动能、大大减少雨滴对表土的溅击。但一些研究结果却与之相反(雷瑞德,1994;周国逸,1997;王彦辉,1986)。
3.3.1 大气降水中雨滴到达林冠面的速度牟金泽等(1983)研究出的雨滴速度计算经验公式一直被研究者采用;钱允祺等(1998)根据Best (1950)提出的雨滴直径分布函数,导出了符合黄土高原降雨雨滴最终速度的分布规律;姚文艺等(2001)根据水力侵蚀模拟方程也提出了计算雨滴速度的经验方程。本研究直接根据雨滴降落过程进行推导。一滴雨滴降落过程中,同时受几种力的作用:重力,它使水滴下落,并在最初阶段作加速运动;空气浮力,它的作用与重力方向相反,一般很小,计算中可忽略不计;压差阻力和粘滞阻力,它们与重力方向相反,正是这种阻力使雨滴的运动速度最终趋向平衡。理论证明(Laiton, 1980)粘滞阻力和压差阻力之和为:

|
(1) |
式中:F1为粘滞阻力和压差阻力之和,k为系数(3.307 5×10-5 kg·s-1m-3),D为雨滴直径,U为雨滴速度,π为圆周率。雨滴的重力为:
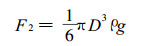
|
(2) |
式中:F2为重力(mg),ρ为雨滴密度,g为重力加速度。下降雨滴达到平衡状态时,即F1=F2,此时的速度为平衡速度,也即雨滴的终速度。根据公式(1)和(2),其大小为:

|
(3) |
式中:Ua为平衡稳定速度。从公式(3)可以看出,给出一个雨滴的直径,就可以求出相应的平衡速度(表 3)。
|
|

现在求出雨滴下降多少高度就可以达到平衡速度。雨滴开始在高空降落时具有势能,在重力作用下,根据牛顿第二定律,其下降的加速度为
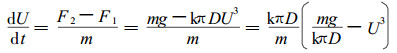
|
(4) |
式中:m为雨滴质量。对上式进行不定积分,用分离变数法求解得:

|
(5) |
式中:t为时间;C为积分常数。上式必须满足初始条件,当t=0时,U=0,于是C为:

|
(6) |
将(6)式代入(5)式,其微分方程的解是:

|
(7) |
利用(3)式给出某雨滴的直径求出相应的平衡速度值代入上式,可得该直径的雨滴从下降开始到达平衡速度所需的时间。然后用所需时间值代入下式(8)积分:

|
(8) |
算出雨滴达到平衡速度时所下降的高度。根据计算,一个直径为7.0 mm的雨滴,下降23 m后,就达到了平衡速度。直径小的雨滴,其高度距离就更小了。因此,有理由认为,大气降水的雨滴远未到达林冠以前就已达到了平衡速度。
3.3.2 冠滴水到达地面的速度大气降水所具有的动能在林冠截留过程中完全消耗在林冠枝叶的阻滞上。冠滴水起初所具有的能量为势能。冠滴水降落到林地的过程是一个由势能转变为动能的过程。可见,冠滴水和大气降水过程的原理是一样的。于是,将公式(7)的U写成时间t的显式并积分可得到:
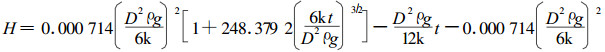
|
(9) |
这样,可用上式,根据杉木林枝下高,算出冠滴水到达林地面所需时间t。把t值代入公式(7),可得到不同直径冠滴水到达林地的速度(表 4)。
|
|

雨滴动能除受速度影响外,还受质量控制。对于一个近似圆球的水滴。只要得知它的直径,就可以求出它的质量。
任何一次降水都是由大小不同的雨滴构成的,随着降雨强度增大,雨滴直径也趋于增大。根据270多次降水资料分析,杉木林外大气降水强度大都为0.5~20 mm·h-1。实测30多次大气降水(雨强0.5~15 mm·h-1)雨滴直径大小与分布表明,当雨强在10 mm·h-1以下,90%的雨滴直径为0.5~3.0 mm。当降雨强度为10~20 mm·h-1时,直径为0.5~3.0 mm的雨滴占总雨滴数的80%以上。根据实测林内降水雨滴直径的结果发现,在观测的雨强范围(0.5~15 mm·h-1)内,无论哪一次林内降水的雨滴直径均大于林外(表 5)。尤其是雨强不大的情况下更为突出。这是因为雨滴降落在叶片表面后在逐渐湿润的表面边缘形成水滴。当重力作用超过张力时,水滴即从边缘落下。可见,冠层枝叶对降水雨滴具有汇聚作用,使得林内降雨中的雨滴直径增大。同时也发现,由于林冠的作用,冠滴水直径受大气降水强度的影响较少。据观测,尽管大气降水强度有所变化,但冠滴水的直径大部分为1.2~3.0 mm,其直径的变化主要受林分郁闭度、冠层厚度、枝叶取向、叶量、叶面积和叶表面对雨水的粘滞性的影响。
|
|
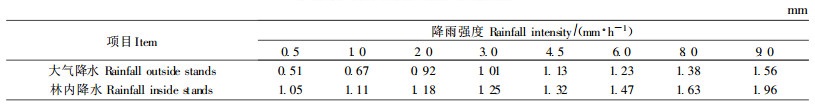
根据不同降水强度的雨滴直径大小和分布数量、雨滴到林冠表面的稳定速度、冠滴水的不同直径分布数量和到达林地的速度,计算了8次不同降雨强度和降水量林冠表面和林地表面承受的动能(表 6)。表 6中林内地面降雨动能包含2个部分,一部分动能是由直接穿透林冠的雨水给予的,它的大小可由林冠表面的总动能乘上系数A(A为1减去郁闭度)得出;另一部分是冠滴水给予的,计算它的方法前面已述。
|
|
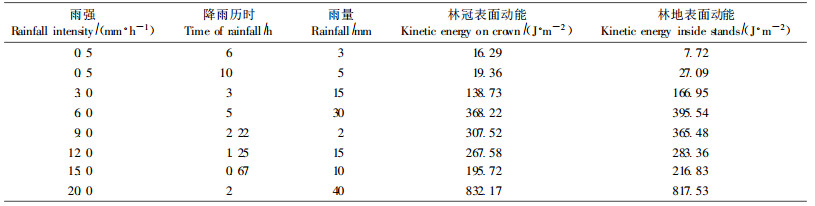
从表 6看出,大气降水经过郁闭度为0.9的林冠截留后,枝下高7 m的杉木林林内降水动能一般都大于林冠上的动能,而且在雨强小的情况下更为突出。这是由于叶对雨水的汇聚使冠滴水的直径增大造成的。实际上,只有2种情况下林冠才能有效地降低降雨动能,一是雨量极少(3 mm以下),林冠层截留了大部分水量;二是雨强极大,直径大的雨滴撞击在枝叶表面时,雨滴动能使雨滴本身击散,直径大的雨滴变成直径小的雨滴进入林内。直径小的雨滴具有的动能较小。
林地土壤被微弱侵蚀的根本原因在于凋落物层对林地的覆盖,森林冠层使林地免受雨水的侵蚀,其作用并不是林冠层对雨水动能的削弱,而是冠层产生凋落物层对林地的保护。
4 结论与讨论杉木林冠层每年吸收的辐射能25.543 0亿J·m-2,占总辐射能82.7%,透过林冠层的辐射量2.530 6亿J·m-2,占总辐射的8.2%,林冠层的反射量2.743 0亿J·m-2,占总辐射的9.1%。林冠层对辐射能的吸收率、透射率和反射率与冠层结构特征因子及枝叶的生长发育状况有关。由于林冠层对辐射能的吸收与反射,减少了到达林下空间的辐射量,改变了系统内能量,造就了林下空间小气候环境。
每年林冠截留水分蒸发耗能6.369 5亿J·m-2,占森林生态系统获取的净辐射能的22.9%,占系统总蒸发散耗能的30.4%。林冠截留水分蒸发耗能使得系统获取的辐射能一部分转化为潜热能,在降雨均匀(雨量和雨强都不大)的月份,林冠截留蒸发耗能在系统能量平衡中占有很重要的位置。
林冠对降水的截留,使大气降水中的雨滴动能消耗在克服枝叶阻力做功上,叶表面汇聚水滴从冠层滴落下来,是一个由势能转变成动能的过程。
杉木林林冠对大气降水雨滴有汇聚作用,使冠滴水水滴直径较林外大。林冠滴水雨滴直径变化和分布范围主要受林分特征因子和叶表面对雨水的粘性的影响,与大气降水强度和降水量关系不大。一般而言,一次性降水中,林冠滴水雨滴动能一般都大于大气降水中雨滴动能,而且在雨强小的情况下更为突出。杉木林冠枝下高度超过7 m,在降水量大于3 mm的情况下,林冠层不能有效地降低降雨动能。只有在雨量极小、林冠层截留大部分水量或是雨强极大、直径大的雨滴在枝叶表面撞击分散的情况下,林冠才能有效地降低降水动能。森林冠层使林地免受雨水的侵蚀,其作用并不是林冠层对雨水动能的削弱,而是冠层产生凋落物层对林地的保护。
陈引珍, 何凡, 张洪江, 等. 2005. 缙云山影响林冠截留因素的初步分析. 中国水土保持科学, 3(3): 318-323. |
蔡丽君, 王国栋, 张仕奇. 2003a. 黄土高原降雨雨滴动能的分布律. 水土保持通报, 23(4): 28-32. |
蔡丽君, 王国栋. 2003b. 风矢量对坡面降雨动能分布的影响. 中国农业大学学报, 8(6): 15-17. |
贡力. 2005. 黄土高原降雨雨滴动能的侵蚀计算. 兰州交通大学学报, 24(4): 43-45. DOI:10.3969/j.issn.1001-4373.2005.04.012 |
江忠善, 宋文经, 李秀英. 1983. 黄土地区天然降雨雨滴特征研究. 中国水土保持, 11(3): 32-36. |
雷瑞德. 1994. 中国森林生态系统定位研究. 哈尔滨: 东北林业大学出版社, 245-251.
|
刘家冈, 张文杰. 1985. 光在林冠中传播. 林业科学, 21(3): 234-240. |
牟金泽. 1983. 雨滴速度计算公式. 中国水土保持, 11(3): 40-42. |
钱允祺, 陶士珩. 1998. 黄土高原降雨雨滴最终速度的分布律. 西北农林大学学报, 26(5): 20-24. |
王万忠. 1987. 黄土地区降雨侵蚀力R指标的研究. 中国水土保持, 15(12): 18-22. |
王彦辉. 1986. 陇东黄土地区刺槐林水土保持效益定量研究. 东北林业大学学报, 8(1): 35-42. |
卫正新, 李树怀. 1997. 不同林地林冠截留降雨特征研究. 中国水土保持, 25(5): 524-530. |
文仕知, 潘维俦, 田大伦, 等. 1989. 杉木人工林生态系统的水文效应. 中南林学院学报, 9(增刊): 29-36. |
谢春华, 关文彬, 吴建安, 等. 2002. 贡嘎山暗针林生态系统林冠截留特征研究. 北京林业大学学报, 24(1): 68-71. |
徐锐. 1983. 关于天然降雨和人工降雨的动能计算方法. 中国水土保持, 11(3): 37-39. |
姚文艺, 汤立群. 2001. 水力侵蚀产沙过程及模拟. 郑州: 黄河水利出版社, 103-109.
|
周光益, 曾庆波, 黄全, 等. 1995. 热带山地雨林林冠对降雨的影响分析. 植物生态学报, 19(3): 201-207. |
周国逸. 1997. 几种常用造林树种冠层对降水动能分配及其生态效益分析. 植物生态学报, 21(3): 250-259. DOI:10.3321/j.issn:1005-264X.1997.03.006 |
周佩华, 豆葆璋, 孙清芳, 等. 1981. 降雨能量的试验研究初报. 水土保持通报, 2(1): 51-60. |
朱劲伟, 崔启武. 1982. 林冠结构和光分布—光的吸收理论的探讨. 林业科学, 18(3): 258-265. |
中野秀章. 1985.森林水文学.李云森, 译.北京: 中国林业出版社, 15-22
|
Best A C. 1950. The size distribution of raindrops. Quarterly Journal of the Royal meteorology, 76(16): 16-36. |
Disrud L A. 1970. Magnitude, probability and effect of kinetic energy of winds associated with rains in Kansas. Kansas Academy of Science, (73): 237-246. |
Kinnell P I A. 1981. Rainfall intensity-kinetic energy relationship for soil loss prediction. Soil Science Society. American Process, (45): 153-155. |
Laiton L. 1980. 生物系统的流体动性. 北京: 科学出版社, 287-290.
|
Salles C, Poesev J, Sempere-Torres D. 2002. Kinetic energy of rain and its functional relationship with intensity. Journal of Hydrology, 257(1-4): 257-271. |
Ulbrich C W, Atlas D. 1985. The effects of drop size distribution truncation on rainfall integral parameters and empirical relations. Journal of Climate and Apply Meteorology, (24): 580-590. |