 2007, Vol. 43
2007, Vol. 43文章信息
- 王永健, 陶建平, 李媛, 余小红, 席一.
- Wang Yongjian, Tao Jianping, Li Yuan, Yu Xiaohong, Xi Yi.
- 华西箭竹对卧龙亚高山森林不同演替阶段物种多样性与乔木更新的影响
- Effects of Fargesia nitida on Species Diversity and Trees Regeneration in Different Forest Cycles of Subalpine Forest in Wolong Nature Reserve
- 林业科学, 2007, 43(2): 1-7.
- Scientia Silvae Sinicae, 2007, 43(2): 1-7.
-
文章历史
- 收稿日期:2006-03-31
-
作者相关文章



自然干扰与斑块动态研究是了解生物群落结构与动态特征的关键(Zang et al., 2005)。植物群落是由具有不同特征的斑块构成的镶嵌复合体(Watt, 1947;Zang et al., 2005),森林循环(forest cycle)理论(Whitmore, 1989;臧润国等,2003)认为任何森林中都存在着由干扰所引起的林分斑块的循环变化过程,在外界干扰因子(风、火、病虫害和树木衰老死亡等)作用下,林分可能形成林隙,林隙干扰驱动了森林的循环更新(Zang et al., 2005)。目前关于热带、温带森林循环更新的研究较多(Emborg, 1998;Zang et al., 2005)。有关川西亚高山暗针叶林林隙特征以及林隙干扰的研究已有报道(杨娟等,2004;王微等,2004),但亚高山森林演替相关研究较少有人涉及,国外已有冷杉林内风灾干扰下的森林演替与更新的相关研究(Nishimura, 2006;Nagel et al., 2006)。根据亚高山暗针叶林演替(刘庆等,2001;吴彦等,2001;赵常明等,2002;郭正刚等,2003;庞学勇等,2005)和林隙更新的相关研究(王微等,2004;杨娟等,2004),并充分借鉴温带和热带森林的斑块动态理论(Watt, 1947)、森林顶级微演替理论(Forcier, 1975)、森林镶嵌循环理论(Emborg, 1998)及森林(发育)循环理论(Whitmore, 1989;臧润国等,2003;Zang et al., 2005),在此提出亚高山暗针叶林循环更新的理论模型,见图 1。认为从阔叶林向针叶林的顶级微演替是亚高山森林循环的主要部分,由外界干扰因子引发的林隙干扰是森林循环的驱动因子,暗针叶林经林隙干扰发展为阔叶林。阔叶林通过树种更新形成针阔混交林,进而发育到暗针叶林阶段。在此过程中,林隙干扰有可能使群落又向阔叶林阶段发展。

|
图 1 亚高山暗针叶林循环演替理论模型 Fig. 1 Conceptual model of forest cycle succession of dark coniferous forest |
林冠与林下物种的相互作用是群落结构和动态的决定因子(Taylor et al., 2004;Griscom et al., 2003)。小径竹(Dwarf bamboo)广泛分布于森林下木层并成为单优种,是影响森林结构动态与演替的关键因子(Noguchi et al., 2004;George et al., 1999)。林下密布的小径竹减弱了林下光的可利用性,影响了林下植物的分布与优势度(Taylor et al., 2004;Holz et al., 2006)。许多研究表明,小径竹对乔木幼苗和林下物种多样性有负面影响(Hiura et al., 1996;Noguchi et al., 2004;Holz et al., 2006)。亚高山森林是本地区物种多样性的库(吴彦等,2001;刘庆等,2001),华西箭竹在卧龙亚高山森林林下分布广泛,箭竹的存在对不同阶段群落微环境可能产生巨大影响,进而影响到个体更新和物种多样性(Suzaki et al., 2005; Noguchi, 2004)。本研究旨在探讨华西箭竹(Fargesia nitida)的存在对森林演替过程中物种多样性与更新的影响。具体解决以下问题:1)华西箭竹存在时,森林循环过程中物种多样性的变化趋势;2)华西箭竹的存在,对各更新阶段森林物种多样性是否产生影响;如有影响,对哪一层次影响最为显著;3)华西箭竹的存在,是否阻碍不同更新阶段森林优势树种的生长与更新。
1 研究区概况研究区位于卧龙自然保护区西南部的四川省林业科学研究院卧龙亚高山暗针叶林生态系统定位站(102°58′21″E,30°51′41″N,海拔2 800 m)附近的亚高山森林。所选区域为典型的岷江冷杉(Abies faxoniana)占优势的暗针叶林,普遍存在着阔叶林与针阔混交林的斑块状分布,华西箭竹为灌木层优势种。卧龙定位站年均气温4.3 ℃,年均相对湿度79%;年均降水量848.9 mm,主要集中于6—9月份,占全年降水的64.25%;年蒸发量772.5 mm,年均日照时数1 185.4 h,土壤为山地棕色暗针叶林土(李振新等,2004)。
2 研究方法 2.1 群落调查2005年8月中旬,正值夏末,植被已经发育成熟,华西箭竹生长已进入稳定期(Li et al., 2002),在卧龙生态站附近的亚高山暗针叶林演替的不同阶段(针叶林-针阔混交林-阔叶林),分别选择有华西箭竹和无华西箭竹的环境设置样地,把华西箭竹密度≥10株·m-2的区域定为箭竹环境(B+),箭竹平均盖度达40%~85%;把华西箭竹密度<2株·m-2且分布较分散的区域定为无竹环境(B-),箭竹平均盖度<2%。选箭竹环境(B+)的针叶林、针阔混交林和阔叶林3个阶段,每个阶段设400 m2的样地4个;选无竹环境(B-)的针叶林、针阔混交林和阔叶林3个阶段,每个阶段设400 m2的样地1个。记录每个样地的地形指标、土壤表层特征及枯落物状况,并使用美国LI-COR公司生产的植冠分析仪(LAI-2000)测定林冠空隙度和叶面积指数。每个400 m2的样地都由16个5 m×5 m的样方组成,在每个5 m×5 m的样方中调查所有乔木和灌木的种名、高度、胸径(基径)及冠幅等,并在每个5 m×5 m的样方中随机设置1个典型的1 m×1 m小样方调查草本,记录草本的种名、平均高度和盖度。同时,测定每个样方内灌木层、草本层总盖度及华西箭竹的密度与盖度。
2.2 更新调查在华西箭竹环境与无竹环境中的6个林分中分别设置4个10 m×10 m的样方,箭竹环境的3个林分在每个400 m2的群落学样地中心设1个10 m×10 m的样方,无竹环境的3个林分在每个400 m2的群落学样地设4个10 m×10 m的样方,调查岷江冷杉和糙皮桦(Betula utilis)的更新。两类乔木大小级分为3级(赵常明等,2004),Ⅰ级幼树:树高(H)≥ 0.33 m,且胸径(DBH)<2.5 cm;Ⅱ级小树:2.5 cm ≤ DBH<7.5 cm;Ⅲ级中树:7.5 cm≤ DBH<22.5 cm。逐株测量Ⅱ和Ⅲ级树木的高度和胸径,测Ⅰ级树木的高度和基径。华西箭竹环境与无竹环境6个林分的4个10 m×10 m的样方中,在10 m×10 m的样方的2条对角线上各设置5个1 m×1 m小样方,即每个10 m×10 m的样方内共10个1 m×1 m的小样方,调查所有乔木树种幼苗(H<0.33 m)数量,每个林分共40个1 m×1 m小样方。
2.3 参数计算1) 乔木层重要值=(相对密度+相对高度+相对优势度)/3;灌木层、草本层重要值=(相对高度+相对盖度)/2。
2) Shannon-Wiener多样性指数(H):H=-∑PilnPi;Simpson多样性指数(D):D= 1-∑Pi2。Pielou均匀度指数(E):E=(-∑PilnPi)/lnS。丰富度指数(R):R= S。Pi为物种相对重要值,S为物种总数。
3) 使用Origin7.0软件作图,运用SPSS11.0软件进行t检验和单因素方差分析,使用Duncan多重比较。
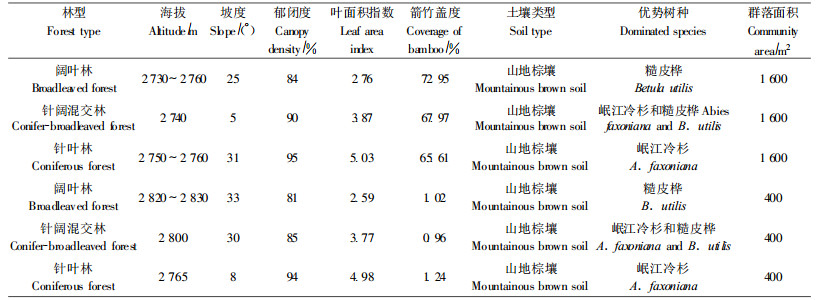
3 结果与分析 3.1 华西箭竹对森林物种多样性的影响 3.1.1 森林演替过程中的多样性动态从表 1中可以看出,存在华西箭竹的3个阶段,随着演替发展,森林叶面积指数增大,更加郁闭,但不同林分间覆盖度差异不大,都处于较郁闭的状态。华西箭竹存在时,演替过程中物种多样性变化见图 2。通过3个演替阶段华西箭竹环境中分别调查的1 600 m2(共4 800 m2)样方各指数的单因素方差分析与多重比较,在乔木层,阔叶林丰富度、多样性和均匀度最高,都显著高于针叶林,针叶林最低;随着演替进程,各指数基本表现为下降趋势。灌木层各指数以针叶林为最高,针阔混交林最低,阔叶林与针叶林各指数间差异不显著,总体表现为随着演替先降低后升高。3类群落草本层各指数间均无明显差异。
|
|

|
图 2 华西箭竹存在时森林循环演替过程中多样性动态 Fig. 2 Diversity dynamics in forest cycle succession with Fargesia nitida H: Shannon-Wiener多样性指数;D: Simpson指数;E:均匀度指数;S:物种丰富度指数。不同字母a, b, c表示差异性显著; ns:差异性不显著(P<0.05)。下同。 H: Shannon-Wiener diversity index; D: Simpson index; E: Evenness index; S: Richness index. Different letter(a, b and c) mean significant difference; ns:Not significant (P < 0.05). The same below. |
3个演替阶段中,除针叶林无竹环境的乔木层物种丰富度低于华西箭竹环境外,所有3层SW多样性指数与丰富度指数都表现为无竹环境高于华西箭竹环境;针叶林乔木层无竹环境的多样性与均匀度指数高于有竹环境,其他乔木层Simpson多样性指数与Pielon均匀度指数在无竹与华西箭竹环境间差异不大;灌木层各指数都表现为无竹环境高于华西箭竹环境;草本层无竹环境的Simpson多样性指数、均匀度指数和华西箭竹环境差异不大(图 3)。

|
图 3 3个演替阶段箭竹环境与无竹环境各层多样性特征 Fig. 3 Diversity characteristics of three layers in B\++ and B\+- conditions of three stages |
3个演替阶段的林分中华西箭竹与无竹环境的群落特征见表 2。地被层特征表明,阔叶林与针叶林下,华西箭竹环境的枯落物和腐殖质平均厚度明显高于无竹环境,且针叶林下华西箭竹环境腐殖质厚度最大,苔藓平均厚度则显著低于无竹环境;针阔混交林下,枯落物、腐殖质和苔藓的平均厚度在2个环境中差异不显著。群落各层特征表明,3个阶段华西箭竹环境的灌木层平均盖度明显高于无竹环境,而华西箭竹环境的草本层盖度显著小于无竹环境。乔木幼苗更新方面,除针叶林下2种环境川滇长尾槭(Acer caudatum)幼苗数量无差异外,3个阶段无竹环境的糙皮桦、川滇长尾槭和冷杉幼苗的数量都明显高于华西箭竹环境。
|
|
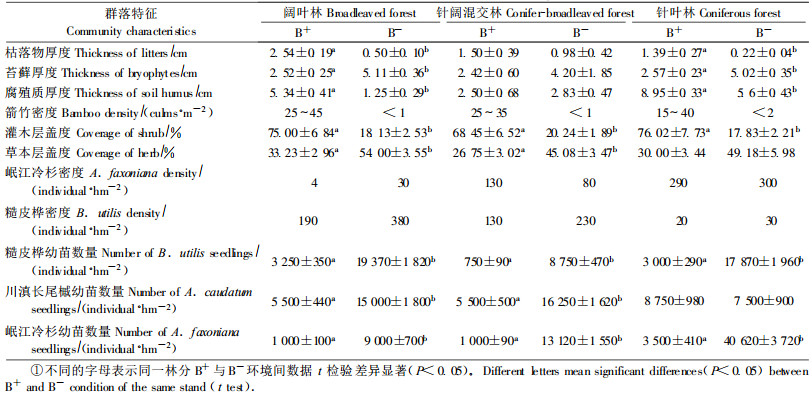
从表 3可以看出,阔叶林下,无竹环境的糙皮桦Ⅰ、Ⅱ级数量明显大于华西箭竹环境,Ⅲ级虽然差异不显著,但也表现出略高于华西箭竹环境的趋势(p=0.055);岷江冷杉各级在无竹与华西箭竹环境均无差异。针阔混交林,无竹环境下糙皮桦Ⅱ级数量显著高于华西箭竹环境,Ⅲ级也有此趋势(p=0.051);华西箭竹环境几乎无Ⅰ、Ⅱ级岷江冷杉,无竹环境此2个径级明显多于华西箭竹环境。针叶林下,无竹环境的糙皮桦Ⅰ级数量显著高于华西箭竹环境;岷江冷杉3个径级的数量都表现了无竹环境显著高于华西箭竹环境。阔叶林无竹环境糙皮桦更新较好,各径级数量最多;针叶林无竹环境冷杉更新最好。
|
|
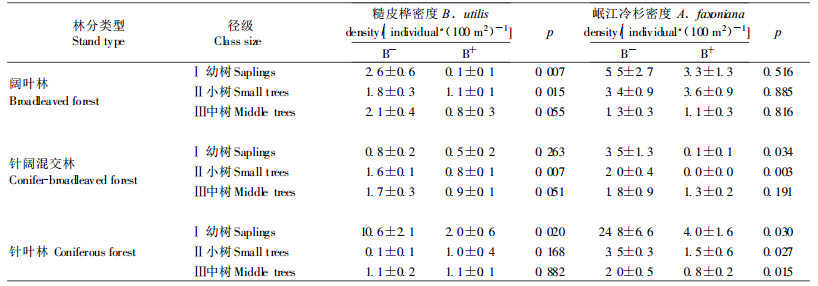
高度/胸径反映了物种对光环境的响应(Emborg et al., 1998)。阔叶林中,华西箭竹环境的岷江冷杉Ⅱ级高度/胸径值大于无竹环境;而糙皮桦Ⅱ级高度/胸径值表现出相反的趋势。针阔混交林中,由于华西箭竹环境的Ⅰ、Ⅱ级冷杉几乎没有,无法准确比较;Ⅱ级糙皮桦高度/胸径值同样表现为华西箭竹环境小于无竹环境。针叶林中,无竹环境的Ⅲ级冷杉高度/胸径值大于华西箭竹环境;Ⅱ级糙皮桦在2个环境中数量很少,无法进行比较(图 4)。

|
图 4 不同阶段存在箭竹与无竹环境乔木高生长 Fig. 4 Height growth of main tree species in B+ and B- conditions of three stages A:阔叶林Broadleaved forest; B:针阔混交林Conifer-broadleaved forest; C:针叶林Coniferous forest. |
小径竹在下木层的繁盛长期影响着森林更新与发育(Taylor et al., 2004)。当华西箭竹存在时,乔木层多样性表现为:阔叶林大于针叶林(Li et al., 2002;郭正刚等,2003;吴彦等,2001),说明华西箭竹的存在并未改变森林循环演替过程中多样性变化总体趋势。3个阶段无竹环境的多样性略高于箭竹环境,阔叶林与针阔混交林无竹环境的物种丰富度高于华西箭竹环境,而针叶林下两类环境丰富度差异不大(图 3),说明华西箭竹的存在一定程度上降低了乔木层物种丰富度,且对阔叶林内树种的影响大于针叶林。对于灌木层,针叶林多样性高于针阔混交林,表现为先降低后升高,与无竹环境的演替趋势相同;在日本和中国西南地区,落叶阔叶林在早春和秋季由于郁闭度很小,利于林下箭竹生长,林下箭竹更高更密(Suzaki et al., 2005),对灌木层物种的生存有很大影响;相反,常绿针叶林终年郁闭的林冠阻碍了小径竹的生长(Reid et al., 1991;Taylor et al., 2004);3个阶段无竹环境与有竹环境的多样性差异明显,说明华西箭竹对灌木层的多样性影响显著,这与许多研究结论相似(Noguchi et al., 2004;Holz et al., 2006),是由于华西箭竹具有盘根错节的地下茎并减弱林下光的可利用性从而阻碍林下灌木的生长与生存(Griscom et al., 2003)。3个演替阶段的有竹环境草本层的多样性无显著差异。
4.2 华西箭竹对树木生长与更新的影响许多森林的组成与结构常常由林下幼树、幼苗库决定(Taylor et al., 2004;Nakashizuka, 1987)。本研究中,3个阶段主要乔木幼苗基本表现为无竹环境多于箭竹环境,无竹环境冷杉幼苗多于糙皮桦(Taylor et al., 2004)。华西箭竹环境低光照及厚厚的枯落物层(表 2)阻碍了幼苗的萌发与生长,川滇长尾槭和糙皮桦幼苗更新对光的需求比冷杉幼苗更高,(Takahashi et al., 2003),华西箭竹的存在增大了它们的死亡率。
树木更新表明:阔叶林下无竹环境更有利于糙皮桦3个径级树木的更新,岷江冷杉3个径级的数量都多于糙皮桦(尤其是华西箭竹环境),冷杉很可能取代糙皮桦成为优势种。华西箭竹环境不利于糙皮桦在针阔混交林进入乔木上层;而针阔混交林内华西箭竹严重影响冷杉更新,但对大径级冷杉的影响很小。针叶林中,对于不耐荫的糙皮桦而言,华西箭竹的存在与否,糙皮桦都无法更新成功,华西箭竹的存在加速了其更新的失败;针叶林下无竹环境更有利于冷杉3个径级树木的更新,华西箭竹的存在阻碍了冷杉的更新进程。
3个阶段有竹与无竹环境中乔木高度径级比(即高生长)表明,阔叶林与针阔混交林下,无竹环境糙皮桦Ⅱ级比华西箭竹环境高,华西箭竹能在其一个生长周期内50年左右对快速生长型落叶乔木糙皮桦Ⅱ级小树产生影响(Taylor et al., 2004)。华西箭竹在其生长周期内主要影响冷杉幼树,但对冷杉幼树高生长影响较小。华西箭竹对于小径级乔木幼苗幼树的影响是明显的,乔木幼树与华西箭竹的“更新生态位”(Emborg et al., 1998)冲突引起它们之间的资源与空间竞争;而其对于较大径级树木,一方面华西箭竹通过降低幼苗幼树更新成功率同时增加幼树更新死亡率(Taylor et al., 2004),另一方面通过与较大径级树木竞争资源(如土壤水分)降低大树生长率,进而影响大树的更新(Griscom et al., 2003;Takahashi et al., 2003)。因此小径竹通过抑制树木更新从而影响了乔木种群动态和森林发育(Nakashizuka, 1987;Taylor et al., 2004)。
与冷杉相比,华西箭竹对糙皮桦更新与生长的影响更为显著,箭竹的存在很大程度上决定了糙皮桦更新必将失败,这有利于加速森林循环演替的进程;但华西箭竹同时减缓了岷江冷杉的更新速率,华西箭竹密度可能是森林演替进程平衡的决定因子,因此,需要进一步研究华西箭竹密度对森林演替的影响。
郭正刚, 刘慧霞, 孙学刚, 等. 2003. 白龙江上游地区森林植物群落物种多样性的研究. 植物生态学报, 27(3): 388-395. DOI:10.3321/j.issn:1005-264X.2003.03.015 |
李振新, 郑华, 欧阳志云, 等. 2004. 岷江冷杉针叶林下穿透雨空间分布特征. 生态学报, 24(5): 1015-1021. DOI:10.3321/j.issn:1000-0933.2004.05.023 |
刘庆, 吴彦, 何海. 2001. 中国西南亚高山针叶林的生态学问题. 世界科技研究与发展, 23(2): 63-69. DOI:10.3969/j.issn.1006-6055.2001.02.016 |
庞学勇, 包维楷, 张咏梅. 2005. 青藏高原东部暗针叶林采伐迹地小气候及植被演替. 世界科技研究与发展, 27(3): 47-53. DOI:10.3969/j.issn.1006-6055.2005.03.010 |
王微, 陶建平, 李宗峰, 等. 2004. 卧龙自然保护区亚高山暗针叶林林隙特征研究. 应用生态学报, 15(11): 1989-1993. DOI:10.3321/j.issn:1001-9332.2004.11.001 |
吴彦, 刘庆, 乔永康, 等. 2001. 亚高山针叶林不同恢复阶段群落物种多样性变化及其对土壤理化性质的影响. 植物生态学报, 25(6): 648-655. DOI:10.3321/j.issn:1005-264X.2001.06.002 |
杨娟, 刘丽娟, 葛剑平, 等. 2004. 卧龙自然保护区林隙干扰特征. 植物生态学报, 28(5): 723-726. DOI:10.3321/j.issn:1005-264X.2004.05.019 |
臧润国, 杨彦承, 林瑞昌, 等. 2003. 海南霸王岭热带山地雨林森林循环与群落特征研究. 林业科学, 39(5): 1-9. DOI:10.3321/j.issn:1001-7488.2003.05.001 |
赵常明, 陈庆恒, 乔永康, 等. 2002. 青藏东缘岷江上游亚高山针叶林人工恢复过程中物种多样性动态. 植物生态学报, 26(增刊): 20-29. |
赵常明, 陈庆恒, 乔永康, 等. 2004. 青藏高原东缘岷江冷杉天然群落的种群结构和空间分布格局. 植物生态学报, 28(3): 341-350. DOI:10.3321/j.issn:1005-264X.2004.03.009 |
Emborg J. 1998. Understorey light conditions and regeneration with respect to the structural dynamics of a near-natural temperate deciduous forest in Denmark. Forest Ecology and Management, 106: 83-95. DOI:10.1016/S0378-1127(97)00299-5 |
Forcier L K. 1975. Reproductive strategies and the co-occurrence of climax tree species. Science, 189: 808-810. DOI:10.1126/science.189.4205.808 |
George L O, Bazzaz F. 1999. The fern understory as an ecological filter: growth and survival of canopy tree seedlings. Ecology, 80: 846-856. DOI:10.1890/0012-9658(1999)080[0846:TFUAAE]2.0.CO;2 |
Griscom B W, Ashton P M. 2003. Bamboo control of forest succession: Guadua sarcocarpa in Southeastern Peru. Forest Ecology and Management, 175: 445-454. DOI:10.1016/S0378-1127(02)00214-1 |
Hiura T, Sano J, Konno Y. 1996. Age structure and response to fine-scale disturbance of Abies sachalinesis, Picea glehnii, and Betula ermanii growing under the influence of a dwarf bamboo understory in northern Japan. Canadian Journal of Forest Research, 26: 289-297. DOI:10.1139/x26-032 |
Holz C A, Veblen T T. 2006. Tree regeneration responses to Chusquea montana bamboo die-off in a subalpine Nothofagus forest in the southern Andes. Journal of Vegetation Science, 17: 19-28. |
Li Z H, Denich M. 2002. Elevational diversity of arrow bamboo(Fargesia spathacea)communities on Mount Shennongjia in Central China. Journal of Forestry Research, 13(3): 171-176. DOI:10.1007/BF02871692 |
Nagel T A, Svoboda M, Diaci J. 2006. Regeneration patterns after intermediate wind disturbance in an old-growth Fagus Abies forest in southeastern Slovenia. Forest Ecology and Management, 226: 268-278. DOI:10.1016/j.foreco.2006.01.039 |
Nakashizuka T. 1987. Regeneration dynamics of beech forest in Japan. Vegetation, 69: 169-175. DOI:10.1007/BF00038698 |
Nishimura T B. 2006. Successional replacement mediated by frequency and severity of wind and snow disturbances in a Picea-Abies forest. Journal of Vegetation Science, 17: 57-64. |
Noguchi M, Yoshida T. 2004. Tree regeneration in partially cut conifer-hardwood mixed forests in northern Japan: roles of establishment substrate and dwarf bamboo. Forest Ecology and Management, 190: 335-344. DOI:10.1016/j.foreco.2003.10.024 |
Noguchi M, Yoshida T. 2005. Factors influencing the distribution of two co-occurring dwarf bamboo species (Sasa kurilensis and S. senanensis) in a conifer-broadleaved mixed stand in northern Hokkaido. Ecological Research, 20: 25-30. DOI:10.1007/s11284-004-0009-6 |
Reid D G, Taylor A H, Hu J, et al. 1991. Environmental influences on Bashania fangiana bamboo growth and implications for giant panda conservation. Journal of Applied Ecology, 28: 855-868. DOI:10.2307/2404212 |
Suzaki T, Kume A, Ino Y. 2005. Effects of slope and canopy trees on light conditions and biomass of dwarf bamboo under a coppice canopy. Journal of Forest Research, 10: 151-156. DOI:10.1007/s10310-004-0123-x |
Takahashi K, Uemura S, Suzuki J, et al. 2003. Understory dwarf bamboo on soil water and the growth of overstory trees in a dense secondary Betula ermanii forest, northern Japan. Ecological Research, 18: 767-774. |
Taylor A H, Huang J Y, Zhou S Q. 2004. Canopy tree development and undergrowth bamboo dynamics in old-growth Abies Betula forests in southwestern China: a 12-year study. Forest Ecology and Management, 200: 347-360. DOI:10.1016/j.foreco.2004.07.007 |
Watt A S. 1947. Pattern and process in the plant community. Journal of Ecology, 35(1): 1-17. |
Whitmore T C. 1989. Changes over twenty-one years in the Kolombangara rain forests. Journal of Ecology, 77: 469-483. DOI:10.2307/2260763 |
Zang R G, Tao J P, Li C Y. 2005. Within community patch dynamics in a tropical montane rain forest of Hainan Island, South China. Acta Oecologica, 28: 39-48. DOI:10.1016/j.actao.2005.02.001 |