 2007, Vol. 43
2007, Vol. 43文章信息
- 江泽慧, 吕文华, 费本华, 任海青, 吴玉章, 王朝晖.
- Jiang Zehui, Lü Wenhua, Fei Benhua, Ren Haiqing, Wu Yuzhang, Wang Zhaohui.
- 3种华南商用藤材的解剖特性
- Anatomical Characteristics of Three Commercial Rattan Canes in South China
- 林业科学, 2007, 43(1): 121-126.
- Scientia Silvae Sinicae, 2007, 43(1): 121-126.
-
文章历史
- 收稿日期:2006-02-05
-
作者相关文章


藤材的解剖结构与藤材性能及其加工利用密切相关。研究藤材的基本解剖特性,对于藤材的归类、开发利用和加工改性等具有重要的指导作用。藤材的商业化取决于以下因素:可及性、直径、节间长度、颜色以及加工特性,这些都基于藤材的基本特性,主要是解剖构造(Walter, 1996)。商业藤材一般具有以下特征(Walter, 1996):1)藤茎的维管束分布均衡,在横截面上,纤维占20%~25%,输导细胞占15%,基本薄壁细胞占30%~35%;2)纤维帽尺寸相等,纤维长度一致,多层结构细胞壁;3)基本薄壁组织细胞小而且壁厚多层。维管束分布密度及纤维成分与输导成分的比例, 纹孔、导管孔径的大小,对藤茎的坚韧程度影响都很大,纤维含量可作为藤茎坚硬度的指标,至于韧性还得考虑到纤维细胞的数量与输导组织成分的比例状况,特别是大型导管孔径的大小等,这些都是选择优良品种的主要依据(腰希申等,1998)。纤维细胞壁的厚度、纤维比率和后生木质部导管分子的直径对藤材密度、初含水率和体积干缩率有着重要影响(Bhat, 1991)。藤种、藤龄、藤茎上的位置、纤维(厚壁组织)比率、密度和含水率是影响强度的重要因子(Bhat, 1992)。本文对我国华南地区的主要用材藤种单叶省藤(Calamus simplicifolius)、白藤(Calamus tetradactylus)和黄藤(Daemonorops margaritae)的纤维长宽、导管分子长宽、组织比量(薄壁细胞、导管、纤维)、单位面积维管束分布密度、后生木质部大导管的大小、维管束的大小、细胞壁厚(薄壁细胞、纤维)、微纤丝角以及纤维素结晶度等解剖特性进行了研究。
1 材料与方法 1.1 材料单叶省藤、白藤和黄藤采自广东省高要市金鸡坑林场。白藤丛生,黄藤和单叶省藤攀生在杉木(Cunninghamia lanceolata)上,约10年生。单叶省藤藤条直径12~15 mm,长10~15 m,节间长度25~35 cm。白藤藤条直径5~8 mm,长4~6 m,节间长度10~15 cm。黄藤藤条直径10~14 mm,长10~15 m,节间长度15~25 cm。藤采收后去除叶鞘,气干。
1.2 试验方法单叶省藤、白藤、黄藤气干藤材各5株,从藤茎梢部往下纵向3等分,记为上、中和下部,截取各部中间藤节的中部和近节部,再按横向内部(近藤芯)、中部和外部(近藤皮)取材,常规制片、离析,显微观测,各指标计测50个值作统计平均。以藤茎中部藤节纤维测微纤丝角和纤维素结晶度,微纤丝角采用偏光显微镜法测定。纤维素结晶度采用红外光谱法测定,用沃康诺指数O'KI或纳耳森·沃康诺指数N·O'KI表示(N·O'KI=a1 372 cm-1/a2 900 cm-1,O'KI=a1 429 cm-1/a893 cm-1,a为谱带强度)。
2 结果与分析 2.1 纤维长度和宽度3种藤材的纤维长度和宽度测试结果(表 1)表明:单叶省藤纤维最长,白藤次之,黄藤最短;单叶省藤纤维最宽,黄藤次之,白藤最窄。与藤节中部纤维相比较,近节处纤维宽度稍窄,长度短10%~20%。自藤芯向藤皮,纤维长度增大,纤维宽度减小。白藤近藤皮外部纤维长度几乎是近藤芯内部的2倍。经F值检验,白藤纤维较短,但与单叶省藤相差不大;黄藤纤维最短,与单叶省藤和白藤都有较显著差异。黄藤纤维较窄,但与单叶省藤相差不大;白藤最窄,与单叶省藤差异显著。对于不同藤种,纤维尺寸在横向的变异情况相同,在纵向稍有差异。表 2是3种藤纤维在纵横向变异的F检验结果,3种藤的纤维尺寸都在纵向(上、中和下部)差异不显著;在横向(内、中和外部)差异很显著;藤节中部与近节处纤维,除在纵向长度差异显著外,无明显差异。
|
|
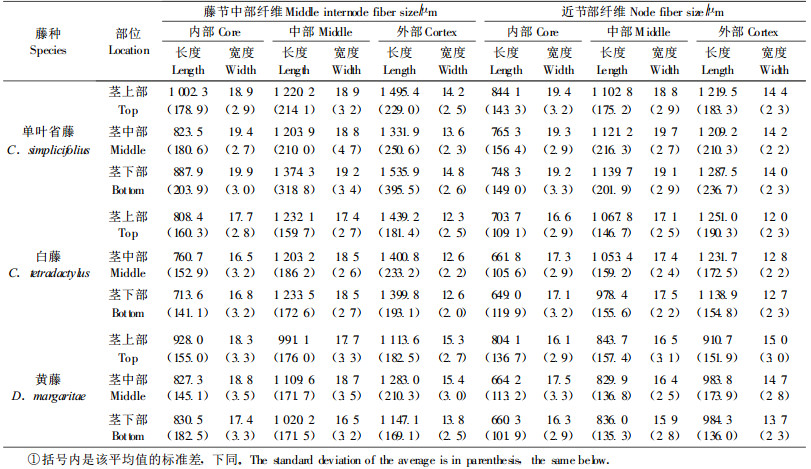
|
|
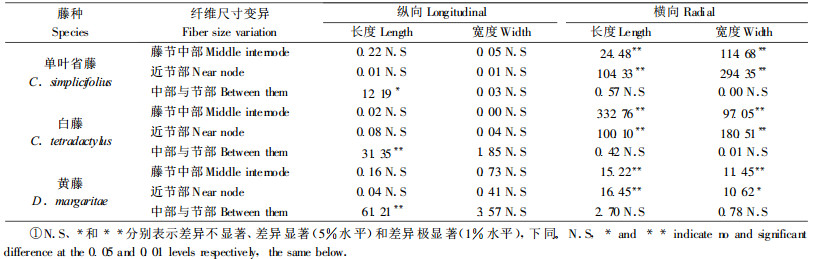
3种藤材的导管分子长度和宽度测试结果见表 3。导管分子长度自藤芯向藤皮呈减小趋势。单叶省藤与白藤的导管分子较长,黄藤导管分子较短,相差近20%。导管分子宽度自藤芯向藤皮减小,近藤皮处不及近藤芯处的一半;单叶省藤最宽,黄藤次之,白藤最窄。藤节中部的导管分子比近节部的导管分子窄,短约20%~30%。经F值检验,单叶省藤与白藤的导管分子长度差异不明显,但黄藤与两者有较显著差异;3种藤材的导管分子宽度存在十分显著的差异。不同藤种导管分子的变异情况,在横向相差不大,在纵向有很大不同。
|
|

表 4是3种藤材的导管分子在纵横向变异的F检验结果。单叶省藤导管分子尺寸在纵向(上、中和下部)无明显差异,在横向(内、中和外部)差异显著;其藤节中部与近节处导管分子相比较,除在纵向长度差异显著外,无明显差异。白藤导管分子在纵向无明显差异,在横向差异显著,但近节处导管分子长度在横向变异小;其藤节中部与近节处导管分子差异显著,但在横向宽度差异不明显。黄藤导管分子长度在纵向变异显著,在横向变异很小;其宽度相反,在纵向变异很小,在横向变异显著;其藤节中部与近节处导管分子,除在横向长度差异显著外,无明显差异。近节处导管分子短,这可能是节部输导性较差原因之一。
|
|
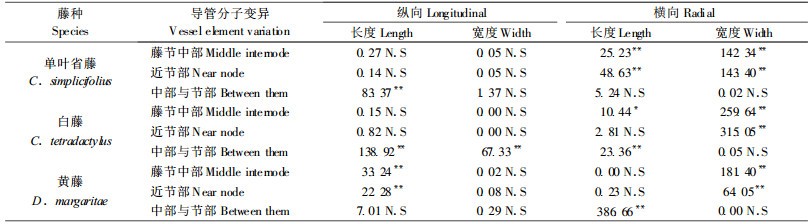
胞壁厚度在某种程度上表征着细胞壁实质含量,胞壁厚度越大,胞壁实质越多,机械力学等性能越好。纤维胞壁厚度是决定藤材物理特性最重要的解剖因子(Bhat et al., 1993)。藤材用于家具制作,可以细胞壁厚度作为强度指标(Walter, 1996)。图 1是3种藤材的纤维细胞和基本薄壁细胞的双壁厚度在横向的变化,2种胞壁厚度在藤茎横截面上的变化一致。单叶省藤的胞壁厚度最大,白藤最小,但白藤和黄藤相差不大。白藤和黄藤的胞壁厚度在藤茎横向自外向内减小。单叶省藤的胞壁厚度在藤茎横向变化很小,在整个横截面上较为一致,表明该藤茎内外性能差异较小。

|
图 1 3种藤材的纤维和基本薄壁细胞双壁厚度的横向变化 Fig. 1 The fiber and parenchyma bi-wall thicknesses of the canes |
图 2是单叶省藤、白藤和黄藤的薄壁细胞、导管和纤维比量在藤茎横向的变化。各组织比量在藤茎横向变化明显,尤其是近藤皮外部与内部和中部的差异较大。薄壁细胞和导管比量自外向内增大,纤维比量自外向内减少,外部纤维含量大,差异明显。纤维比量在一定程度上表征藤茎的力学强度;纤维细胞的数量与输导组织成分的比例状况对藤材韧性有重要影响(腰希申等,1998)。藤材的塑性变形受薄壁细胞含量影响;藤材的抗拉、抗压强度和弹性模量与其密度或纤维比量显著正相关,与其薄壁组织比量显著负相关(蔡则谟等,2003)。近藤皮处的纤维比量最大,薄壁细胞和导管比量最小,使得藤茎外部比藤芯更坚韧。白藤薄壁细胞含量高,内外差异小,因而白藤柔韧工艺特性好。纤维比量的变化率和变化梯度反映藤材质量;下降率小,下降梯度平缓,为材质良好的构造特征。单叶省藤的纤维比量最大,且内外差异较小,因而单叶省藤材质最好,综合利用价值最高。

|
图 2 3种藤材的薄壁细胞、导管和纤维比量 Fig. 2 The basic parenchyma, vessel and fiber ratios of the canes |
维管束分布密度对藤茎坚韧程度有重要影响(蔡则谟等,2003)。表 5是3种藤材的维管束大小和分布密度在藤茎横向的变化。维管束分布密度在横向差异显著,近藤皮分布多,近藤芯分布少,但中部和内部的差异不显著,藤茎外部比藤芯坚韧。单叶省藤的维管束分布密度最小。黄藤的维管束分布密度在横向差异十分显著。维管束分布密度与维管束大小正相关。维管束在藤茎横向自外向内增大,近藤皮外部的维管束小,芯部和中部的维管束大。维管束长度和宽度的变化一致。单叶省藤的维管束大,黄藤较小,白藤最小。维管束分布和大小的横向变化,单叶省藤差异小,黄藤大,白藤居中。结构上的差异决定性能上的差异,因此,单叶省藤材质均匀,白藤稍差,黄藤横向材质差异最大。
|
|
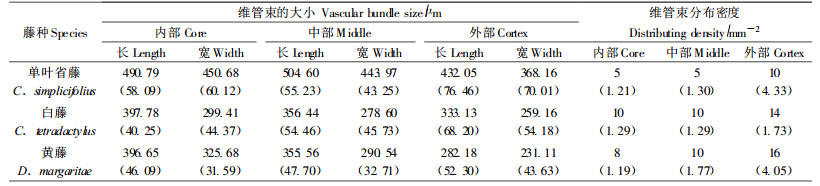
后生木质部导管分子直径对藤茎的坚韧程度影响较大,特别是大型导管分子孔径的大小对韧性有重要影响(腰希申等,1998)。由图 3,3种藤材的后生木质部大导管分子直径在横截面上自外向内增大,变化明显。藤茎外部大导管分子较小,芯部和中部大导管分子较大。单叶省藤的后生木质部大导管分子最大,黄藤较小,白藤最小。芯部和中部大导管分子的差异,单叶省藤不明显,白藤和黄藤差异较明显,中部较大,芯部最大。

|
图 3 3种藤材的后生木质部大导管直径 Fig. 3 The metaxylem vessel diameters of canes |
纤维微纤丝角为细胞次生壁中层S2层微纤丝排列方向与细胞主轴所形成的夹角,它对木材的物理、力学和化学性能都有很大影响,是木材机械性能的主要决定因子,特别影响木材的弹性模量和异向收缩(Walke et al., 1995; Cave et al., 1994),与木材强度和硬度等密切相关(洑香香等,2002)。纤维微纤丝角对藤材性能的影响,与对木材性能的影响相似,藤的力学性质和干缩特性等与之密切相关(Abasolo et al., 2000)。表 6是3种藤材纤维微纤丝角在藤茎横向以及藤节中部和节部的变化。微纤丝角自藤皮向藤芯呈增大趋势,这是机械组织分布适应外界作用力的结果(蔡则谟,1989)。近节部纤维的微纤丝角明显比藤节中部纤维的微纤丝角大,因此,藤节往往是藤破坏时的脆弱点。黄藤纤维的微纤丝角变异大,表明黄藤的力学等相关性质变异大。
|
|

纤维素结晶度是纤维素构成的结晶区占纤维素整体的百分数,表征纤维素聚集态结晶程度,在一定程度上反映纤维的物理和化学性质,是评价生物质材料力学性质和纤维素品质的重要依据。随着结晶度增加,纤维素的断裂强度、弹性模量、硬度、密度、尺寸稳定性等提高;但延伸性、吸湿性、润胀性、吸附性、韧性、柔软性和化学反应性等降低(吴顺昭等,1991;秦特夫等,2005)。纤维素结晶度对藤材的铸型或重组有重要影响(Abasolo et al., 2003)。表 7是3种藤材的纤维素结晶度,沃康诺指数(O'KI)或纳耳森·沃康诺指数(N·O'KI)越大,表明纤维素结晶度越大。可见,藤材纤维素结晶度明显比木材低,应用于家具工业,可经适当加热后,在不改变机械特性的情况下铸模、弯曲成型。纤维素结晶度的横向变异,白藤较大,单叶省藤次之,黄藤最小。藤芯纤维素结晶度高于藤皮,藤芯机械强度比藤皮好,但藤皮比藤芯更容易弯曲。单叶省藤的结晶度指数最大,黄藤最小,白藤居中,这表明相关材性单叶省藤好于白藤,白藤好于黄藤。
|
|

基于藤材的解剖特性及其种间和茎内差异,进行藤种或藤茎剖分归类,适材适用,可提高藤的利用价值和加工层次。对单叶省藤、白藤和黄藤的解剖研究表明,藤茎横向(内、中、外部)的材质差异较大,藤茎纵向(上、中、下部)的材质差异较小。这表明将藤茎沿纵向圆周剖分,不涉及横向变化,所剖分出的藤条性质是一致的,因而,在实际应用中,将藤茎内外材性差异较大的藤材剖分分类使用。纤维长度、纤维比量、维管束分布密度、纤维胞壁厚度等自藤皮向藤芯减小,藤皮与藤芯的纤维比量有很大差异,藤皮部的维管束数量高于藤芯部1倍以上。纤维宽度、导管分子长度与宽度、薄壁细胞比量、导管比量、维管束大小、后生木质部导管分子直径和纤维微纤丝角等自藤皮向藤芯增大。藤芯纤维素结晶度比藤皮高。藤茎的维管束分布密度、纤维比量及纤维壁厚率等解剖特性,决定了藤茎外部强度大于藤芯,如果二者差别较小,认为藤茎质地均匀而良好;反之,质量较差(蔡则谟,1994)。
研究结果表明,单叶省藤、白藤和黄藤均具备良好商用藤材的结构特征(Walter, 1996)。单叶省藤的纤维比量、纤维胞壁厚度、纤维素结晶度以及纤维、导管分子、维管束和后生木质部大导管分子的尺寸等最大,薄壁细胞比量和维管束分布密度等最小,而且在藤茎横向的内外差异最小,变化梯度平缓,因此,单叶省藤材质最好。同理,白藤材质次之,黄藤材质稍差。此外,单叶省藤藤径较大,节间长度较大,颜色较浅,从而综合利用价值最大,为我国棕榈藤栽培的首选藤种。综上,单叶省藤,中径藤,藤皮及藤芯材质差异小,质地优良,宜原条或加工成藤皮和藤芯使用,用于家具、织物或手工艺品等。黄藤,中径藤,纤维比量较大,但藤茎横向材质差异显著,藤芯脆弱,质地中等,宜原条或加工成藤皮和藤芯使用,与单叶省藤用途相似。白藤,小茎藤,纤维比量较大,宜原条使用,用于捆扎或制作工艺品、家具和农具等。
藤材导管分子横向变异十分显著,藤芯导管分子长而宽,藤茎外部导管分子短而窄,因而在实际应用中,通过浸渍处理方法改善藤材特性的应用相当受限。藤茎的渗透途径限于轴向,主要是藤茎内部,横向穿透非常弱。研究藤材构造与渗透性的关系,提高藤材的可渗透性,对于藤材的防霉、防腐等改性处理意义重大,是藤材利用的重要研究方向。
蔡则谟. 1989. 四种藤茎维管组织的分布. 植物学报, 31(8): 569-575. |
蔡则谟. 1994. 藤茎的轴向抗拉强度试验. 林业科学, 30(1): 93-95. |
蔡则谟, 许煌灿, 尹光天, 等. 2003. 棕榈藤利用的研究与进展. 林业科学研究, 16(4): 479-487. DOI:10.3321/j.issn:1001-1498.2003.04.018 |
洑香香, 杨文忠, 方升佐. 2002. 木材微纤丝角研究的现状和发展趋势. 南京林业大学学报:自然科学版, 26(6): 83-87. |
秦特夫, 黄洛华. 2005. 5种不同品系相思木材的化学性质Ⅰ.木材化学组成及差异性.. 林业科学研究, 18(2): 191-194. DOI:10.3321/j.issn:1001-1498.2005.02.017 |
吴顺昭, 王义仲. 1991. 黄藤材之物理性质. 中华林学季刊, 24(2): 99-110. |
腰希申, 李旸, 许煌灿, 等. 1998. 棕榈藤的电镜观察Ⅰ.藤基部分的电镜观察. 林业科学, 34(3): 104-109. DOI:10.3321/j.issn:1001-7488.1998.03.015 |
Abasolo W P, Yoshida M, Yamamoto H, et al. 2000. Microfibril angle determination of rattan fibers and its influence on the properties of the cane. Holzforschung, 54(4): 437-442. |
Abasolo W P, Yoshida M, Yamamoto H. 2003. Thermal softening of rattan canes: influence of the hemi cellulose-lignin matrix. World Bamboo and Rattan, 1(4): 32-36. |
Bhat K M. 1991. Mohan verghese anatomical basis for density and shrinkage behaviour of rattan stem. Journal of the Institute of Wood Science, 12(3): 123-130. |
Bhat K M, Thulasidas P K. 1992. Strength properties of ten south Indian canes. Journal of Tropical Forest Science, 5(1): 26-34. |
Bhat K M, Walter L. 1993. Anatomy and identification of south Indian rattans (Calamus species). Journal of IAWA, 14(1): 63-76. DOI:10.1163/22941932-90000578 |
Cave I D, Walker J C F. 1994. Stiffness of wood in fast-grown plantation softwoods:the influence of microfibril angle. Forest Products of Journal, 44(5): 43-48. |
Walke J C F, Bunerfield B G. 1995. The importance of microfibril angle for the processing industries. New Zealand Forestry, 40(4): 34-40. |
Walter L. 1996. Structural research on bamboo and rattan for their wider utilization. Journal of Bamboo Research, 15(2): 1-14. |