 2007, Vol. 43
2007, Vol. 43文章信息
- 刘足根, 朱教君, 袁小兰, 谭辉.
- Liu Zugen, Zhu Jiaojun, Yuan Xiaolan, Tan Hui.
- 辽东山区长白落叶松天然更新调查
- Investigation and Analysis of the Natural Regeneration of Larix olgensis in Mountain Regions of Eastern Liaoning Province, China
- 林业科学, 2007, 43(1): 42-49.
- Scientia Silvae Sinicae, 2007, 43(1): 42-49.
-
文章历史
- 收稿日期:2006-04-11
-
作者相关文章



2. 江西省环境保护科学研究院 南昌 330029;
3. 江西农业大学商学院 南昌 330013
2. Jiangxi Research Institute of Environmental Science and Protection Nanchang 330029;
3. Commercial School of Jiangxi Agriculture University Nanchang 330013
天然更新充分依靠自然的力量来实现森林生态环境的自然恢复,并且能够培育出合乎自然规律的高生物多样性和高生态质量的森林,而其育林成本大约只是人工更新造林成本的1/4,是一种低投入而高产出的森林培育方式(贾云飞等, 2001)。由于这种传统的森林更新方式更适合现代森林可持续性经营及生物多样性保护的原则,因而在国内外备受关注(董广平, 2001; Gong et al., 1999)。
长白落叶松(Larix olgensis)是我国东北地区一种重要的商业用材林树种,在东北地区广泛栽培。过去几十年中,当地人们比较关注经济发展,在林业政策方面,更多的是关心人工林的经济(Zhang et al., 2000),因此,更多的研究主要集中在改善人工林的经济效益,如森林采伐经营和幼苗培育的管理等方面(洪启法等, 1993; Liu, et al., 1998),而对于人工林的生态研究却被忽视了(Li, 2004),尤其是对人工林的天然更新研究。长白落叶松林下长白落叶松天然更新不良(王战等, 1990; Tsuyuzaki, 1994; Okitsu et al., 1995; Liu, 1997),甚至在采伐林地中(Tsuyuzaki, 1994; Okitsu et al., 1995; Liu, 1997)。有研究认为落叶松的天然更新仅出现在大的干扰以后,如火烧、风倒和洪水过后(Tsuyuzaki, 1994; Liu, 1997; 郝占庆等, 2002),但具体为什么出现这种现象并未进一步开展具体研究。林木成功的天然更新一般需要3个基本条件:1)足够的种源;2)适宜萌发的苗床;3)幼苗存活和生长的适宜生境条件(Gong et al., 1999; Zhu et al., 2003b)。本文是“长白落叶松天然更新主要障碍因素研究”的一部分,其中已经确定了种源不是影响长白落叶松天然更新不良的主要因素,且在调查中也发现了许多当年生长白落叶松幼苗存在,然而,许多1年生幼苗在更新循环的早期阶段就消失了。因此,种源和适宜的种子萌发条件也不是限制因素。那么,很可能是长白落叶松幼苗生长和存活的阶段存在障碍,因此这部分将是本文要探讨的主要内容。
1 研究地概况和研究方法 1.1 研究地概况辽东山区为长白山西南延伸部分,境内有浑河、太子河、清河、柴河等主要河流。全区主要基岩为花岗岩、片麻岩、玄武岩和其他变质岩。气候属暖温带大陆性季风气候,冬季漫长寒冷多雪,夏季炎热多雨,年平均气温4.7 ℃,大于10 ℃的年活动积温2 800~3 100 ℃。最冷出现在1月,最热出现在7月,年均降水量810.9 mm,降水量集中在6—8月,无霜期大约130 d,植物生长季在4—9月(Zhu et al., 2006)。土壤多为棕色森林土和暗棕色森林土,全区植被属长白区系,地带性植物群落是阔叶红松(Pinus koraiensis)林,在历经长期的人为和自然干扰后,目前绝大部分已演变为次生林和人工林,人工林总面积1.15×105 hm2,主要以落叶松为主。研究地位于中国科学院沈阳应用生态研究所清原森林生态试验站(N41°51.080′,E124°56.171′,EL564 m)长白落叶松人工试验林内,样地布置在1965年种植的长白落叶松人工林内(表 1)。
|
|
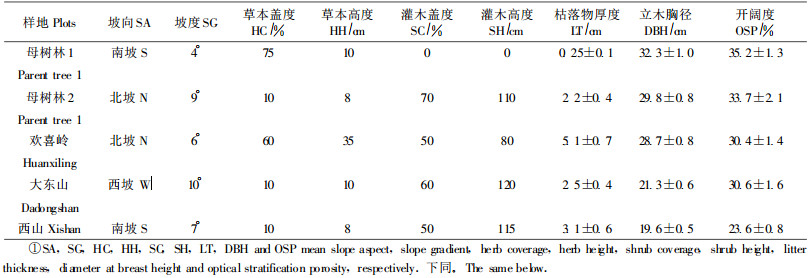
2005年在清原站长白落叶松人工林内,选取了母树林1(P1)、母树林2(P2)、欢喜岭(P3)、大东山(P4)和西山(P5)共5块长白落叶松人工林,在每块样地内选择3个面积为20 m×20 m的固定样地,在每个样地的2条中心连线上,每隔1 m,设置1 m×1 m的小样方(共60个)。从4月至9月,每个月的中旬调查1次长白落叶松天然更新数量和更新频度(幼苗更新频度=更新幼苗出现的样方数/总样方数×100%),在每个小样方内,随机标记5株落叶松幼苗(如下次调查中有幼苗死亡,用同样的方法补充相同数量的幼苗)观察其生长状况,记录其高。在样方内进行幼树调查,如样方内幼树没有或太少,则在整个样地内清查幼树。
除调查以上5块样地外,还通过踏查以及访问当地林场工作人员、居民,重点调查了当地有长白落叶松天然更新幼树出现的地方,包括皆伐地、火烧迹地、采伐迹地和一些样地林缘,并调查记录了这些样地的生境状况。另外,根据曾东等(2000)对新疆落叶松(Larix sibirica)幼树幼苗的定义,本文对长白落叶松幼苗和幼树做以下规定:幼苗高为 < 30 cm,幼树高为≥30 cm。
1.2.2 种子库调查在2004年4月份,在以上5块样地的每个样方内,随机选取了20个30 cm×30 cm的小样方,把小样方内所有的枯落物和0~5 cm的土壤用塑料袋带回实验室计算种子数量。
1.2.3 地被物调查在2004年6月份,记录了每个小样方内的草本和灌木的种类、盖度以及枯枝落叶的厚度。
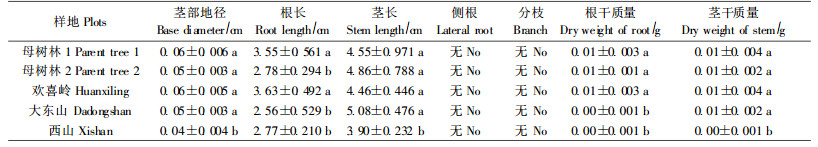
1.2.4 生物量测定在每个样地的最后一次调查更新幼苗时,随机轻轻拔出10株死亡幼苗,观察幼苗根系生长状况。并分离根系和泥土,带回实验室,测定幼苗根部地径、根长和茎长,并把根和茎放在105 ℃的烘箱里烘24 h,测定各部分干质量。
1.2.5 林冠开阔度测定林冠开阔度=1-林分郁闭度(Zhu et al., 2003a)。对每块样地进行立木调查后,在每块样地的每个样方内随机选择了9个点,在黄昏时,用数码照相机(Nikon, coolpix, Japan, f=7~21 mm)和180°鱼眼镜头转换器(Nikon, FC-E8, f=8~24 mm, Japan)距地1 m处,测定林冠开阔度(Zhu et al., 2003a),结果见表 1。
2 结果 2.1 幼苗更新状况在调查的5块样地中,并没有发现长白落叶松幼树,全都是1年生长白落叶松幼苗,且各样地更新状况良好,6月份幼苗更新频度除大东山样地为98.3%外,其他样地更新频度都为100%。从表 1可以看出,各样地由于林分结构不同(林木胸径和林分开阔度不同),地面种子库数量有显著差异(P < 0.05),依次为母树林1 > 母树林2 > 欢喜岭 > 大东山 > 西山,相应的6月份幼苗株数也表现出了相似的结果,即幼苗出现数量也为母树林1 > 母树林2 > 欢喜岭 > 大东山 > 西山,尤其是在母树林1样地中,每m2长白落叶松幼苗在6月份平均达到了79.8株(见表 2)。即使更新幼苗数量最小的西山样地,每m2也达到了11.2株。长白落叶松幼苗萌发的数量受草本和灌木盖度的影响不明显,除了欢喜岭(P3)样地6月份幼苗更新数量与草本盖度呈负相关外(P < 0.05),其他样地6月份幼苗出现的数量与各样地草本和灌木的盖度之间并不存在线形关系(P>0.05)(图 1)。
|
|


|
图 1 各样地草本、灌木与落叶松种子萌发的关系 Fig. 1 Relationship between herbs, shrubs and germination of L. olgensis seeds P1、P2、P3、P4和P5分别为母树林1、母树林2、欢喜岭、大东山和西山样地。P1, P2, P3, P4 and P5 referring to the plots of parent tree 1, parent tree 2, Huanxiling, Dadongshan and Xishan, respectively.下同。The same below. |
各样地幼苗在不同调查时期具有明显的动态变化(图 2)。从图 2可以看出,4月中旬各样地都没有发现长白落叶松幼苗,5月中旬调查时,各样地都长出了落叶松幼苗,且各样地在6月份调查时,幼苗数量达到最多,此后各样地幼苗开始急剧减少,8月份调查时,欢喜岭和西山2个样地已经没有更新幼苗了,到9月份调查时,大东山样地每m2仅为0.1株,而其他样地幼苗保留下来的也很少,母树林1样地保留下来的幼苗为幼苗最多时(6月份)的6.6%,母树林2样地也为最多幼苗时(6月份)的5.1%。另外,各样地幼苗数量之间在每次调查时都差异显著(P < 0.05)。

|
图 2 各样地长白落叶松幼苗在生长季节的更新数量动态 Fig. 2 Dynamic of regeneration number of L. olgensis seedlings at different plots during the growing season 不同字母表示差异显著,P < 0.05. Marked with the different letters were significantly different, P < 0.05.下同。The same below. |
长白落叶松幼苗在林下生长缓慢,5月中旬,也就是幼苗萌芽不久,幼苗单株平均高就为2.5 cm。从5月份到9月份,各样幼苗的平均高生长幅度不超过3 cm,单株幼苗最高出现在母树林样地2,也不超过6 cm(图 3)。从图 4可以看出,幼苗的叶片数量与高生长表现出相同趋势,不过平均叶片数量增长的时间一直到了8月份,此后,各样地幼苗平均叶片数量也开始减少。在这5块样地中,大东山样地幼苗的平均叶片数量一直为最多,但单株幼苗最多叶片数量也不超过25针。经过一个生长季节的幼苗,其器官生长发育缓慢(表 3)。从表 3可以看出,各样地1年生幼苗在生长季节末仍然纤细矮小,各样地幼苗平均地径为0.04~0.06 cm,且没有分枝;根系还没有完全发育,只有主根,侧根还没有形成,而且长度短,平均根长仅为2.56~3.63 cm。当然,相应这些器官的生物干质量也较低(表 3)。

|
图 3 各样地长白落叶松1年生幼苗高生长状况 Fig. 3 Height growth of one-year seedling for L. olgensis at different plots |

|
图 4 各样地1年生长白落叶松幼苗叶片数量 Fig. 4 Leaf number of one-year seedlings for L. olgensis at different plots |
|
|
研究区长白落叶松人工林下幼树很少,只在以下几个样地发现了幼树:
1) 火烧迹地 面积为5 100 m2的黄代沟屯后山(N41°52.335′,E124°56.274′),2000年春季发生火灾,当时火烧强度为中度,火烧使地面的草本和灌木全部死亡,枯枝落叶层遭到破坏,成熟长白落叶松死亡率大约为70%。现在在火烧迹地中没有灌木或者灌木比较矮(< 30 cm)的地方,能发现落叶松幼树,呈簇状分布,其平均高为0.31 m,地径为0.27 cm,但在灌木较多的样地,没有找到长白落叶松幼树。
2) 间伐迹地 面积为960 m2的红松长白落叶松混交林(N41°54.935′,E124°56.278 ′),2000年当地林场为使红松生长得更好,把所有的长白落叶松间伐了。在这块间伐迹地,长白落叶松幼树较多,在林隙中呈片状分布,平均高为0.34 m,平均地径为0.51 cm。
3) 母树林2林缘 母树林2样地北林缘,靠近马路边,长白落叶松幼树较多,且生长良好,其平均高为0.82 m,平均地径为0.73 cm,最高的幼树为4.2 m,地径为6.6 cm。
4) 欢喜岭林缘 欢喜岭东林缘,靠近路边,林缘路两边零散地分布着长白落叶松幼树,其平均高为0.34 m,平均地径为0.52 cm。
另外,从表 4还可以看出,这些样地有些共同点,就是样地都处于北坡,坡度较缓,枯落物少或没有,幼树出现的地方没有或者只有很少的灌木,草本没有或者草本的高度低于落叶松幼树,都受过较大的干扰(如火或采伐)以致林内开阔度较大或者处于林缘地。
|
|

长白落叶松开始萌发的温度约为10 ℃(李贵, 1996)。当地气温通常在4月末5月初上升到10 ℃,此时落叶松种子开始萌发。5块样地中,种子库种子数量为母树林1>母树林2>欢喜岭>大东山>西山,各样地幼苗萌发的数量也为母树林1>母树林2>欢喜岭>大东山>西山,即幼苗出现的数量与种子库种子数量较一致(表 2)。这与幼苗萌发的时间主要由土壤温度和湿度决定,幼苗萌发数量由土壤种子库中种子数量决定的结果一致(杨跃军等, 2001; McLaren et al., 2003)。另外,从图 1可以看出,草本和灌木的盖度与落叶松幼苗出现的数量之间并没有明显的线形关系(P>0.05),也就是说草本和灌木不影响长白落叶松幼苗的萌发,这个结果与Sharew等(1997)发现草本和灌木不影响非洲圆柏(Juniperus procera)幼苗萌发的结果相似。
3.2 长白落叶松幼苗存活和生长从表 2可以看出,2004年6月份,5块样地中1年生长白落叶松生幼苗数量较多,达到11.2~79.8株·m-2,这对更新来说是足够的。然而,2005年5月份调查时,这些样地都没有发现2年生幼苗,这说明了1年生长白落叶松幼苗在林内不能正常存活和生长,也就是不能正常天然更新。林木的天然更新是一个复杂的生态学过程,幼苗的存活和生长依赖于许多微生境因子的影响,如光照条件、草本、灌木、枯落物、水分和养分等(班勇等, 1995; Archibold et al., 2000; Zhu et al., 2003b; 董希斌等, 2003)。当然,林木自身的特性对幼苗的存活和生长也起到了重要的作用(Tao et al., 1987; 曾德慧等, 2002),如树种特性和种粒大小。
长白落叶松是强阳性树种,在幼苗生长期间需要充足的光照才能维持自身的发展,且长白落叶松属于小粒种子,质量轻,呈扁平三角形且带翅,很容易受林内死、活地被物和根系构成的隔离带的影响,这些特征不利于长白落叶松种子直接接触土壤,因而在自然情况下较难得到充足的水分和养分,不利于幼苗的生长。清原实验站长白落叶松人工林大多于20世纪50— 60年代种植,地面已积累了较厚的枯落物;另外,试验地落叶松林已经历了1次或多次的抚育间伐。间伐强度小的,则林分开阔度小,光照不足;间伐强度大的,开阔度大,则林下草本和灌木多。而光照不足或草本灌木和枯落物多的生境都不利于长白落叶松幼苗的生长和存活。从表 3、图 3和图 4可以看出,1年生长白落叶松幼苗的地径、茎干、根长和叶片等生长不良,而且还没有形成分枝和侧根,结果不利于幼苗生长和成活(高贤明等,2003; Facelli et al., 2005)。这与其他研究人员发现光、草本灌木与枯落物影响早期针叶树幼苗生长和存活的结果相似(DeLong et al., 1997; Cornett et al., 1998)。
辽东山区的气候条件不利于长白落叶松幼苗的存活和生长。在辽东山区,7、8月份雨水较集中,使枯落物层积水过多,而长白落叶松幼苗生长纤弱,且根系还没有进入到土壤,这样很容易造成幼苗涝死。接下来的8月末和9月初,连续的高温使得枯落物变得干燥,又容易造成幼苗旱死。进入10月,当地气温开始急剧下降,接下来的霜冻和寒冷很可能是剩下部分幼苗死亡的主要原因。
综合以上分析,影响辽东山区长白落叶松幼苗存活和生长的主要因素是光、草本灌木和地面枯落物。
3.3 长白落叶松幼树出现的生境从以上分析幼苗存活和生长的因素可以看出,长白落叶松幼苗只在光照比较充足,地面死、活地被物没有或较少的生境才能存活和生长。这些条件与长白落叶松幼树出现的生境相似。火烧后,林内开阔度增加,光照增强,而且火烧过程中,地面的枯落物和草本灌木也遭到破坏。这样,火烧后的生境,幼苗的根系容易进入土壤,又有强的光照,使幼苗能正常存活和生长(单建平等, 1990; Schroeder et al., 2002)。一段时间后,火烧迹地的草本灌木重新迅速生长,又使得很多落叶松幼苗缺乏光照以及竞争不过草本灌木而死亡。这样,只在没有或较少的草本灌木的生境还保留了落叶松幼树。皆伐地生境类似于火烧迹地,皆伐后有足够的光照,即使当时有幼苗出现,但随后密集的草本灌木快速生长也使落叶松幼苗缺乏光照而死亡。红松间伐迹地,间伐落叶松后,林中出现了较大的空隙;且红松林下草本灌木很少,这些都为落叶松正常天然更新提供了良好的条件。林缘光照比较充足,但大多林缘草本灌木多、土壤水分过多或过少、瘠薄不利于幼苗生长,因此,只有少数林缘的生境具备形成长白落叶松幼树的条件。如母树林2样地和欢喜岭样地林缘。另外,本研究发现长白落叶松幼树出现的生境都在北坡,这很可能与北坡土壤湿润而水分较多(安守芹等, 1997; Day et al., 2005)有关,这表明长白落叶松幼树出现需要较湿润的生境。
安守芹, 张吉术, 李华. 1997. 兴安落叶松林冠下天然更新的研究. 内蒙古林学院学报:自然科学版, 19(1): 1-8. |
班勇, 徐化成. 1995. 兴安落叶松老龄林分幼苗天然更新及微生境特点. 林业科学研究, 8(6): 660-664. |
单建平, 徐振邦, 陶大立. 1990. 试论火与兴安落叶松种子发芽条件的关系. 林业科学, 26(6): 545-549. |
董广平. 2001. 日本的天然林管理和天然林更新. 世界林业研究, 14(4): 457-464. |
董希斌, 王立海. 2003. 采伐强度对林分蓄积生长量与更新影响的研究. 林业科学, 39(6): 122-125. DOI:10.3321/j.issn:1001-7488.2003.06.020 |
高贤明, 杜晓军, 王中磊. 2003. 北京东灵山区两种生境条件下辽东栎幼苗补充与建立的比较研究. 植物生态学报, 27(3): 404-411. DOI:10.3321/j.issn:1005-264X.2003.03.017 |
郝占庆, 郭水良, 曹同. 2002. 长白山植物多样性及其格局. 沈阳: 辽宁科学技术出版社, 126-127.
|
洪启法, 张凡, 刘健华. 1993. 长白落叶松苗最佳密度的小气候研究. 吉林林学院学报, 9(4): 55-61. |
贾云飞, 阳艳兰. 2002. 浅谈小兴安岭林区的森林促进天然更新问题. 林业勘查设计: 11-12. |
李贵. 1996. 长白落叶松播种育苗丰产经验. 林业科技通讯: 26-27. |
王战, 张颂云. 1990. 中国落叶松林. 北京: 中国林业出版社, 77-78.
|
杨跃军, 孙向阳, 王保平. 2001. 森林土壤种子库与天然更新. 应用生态学报, 12(12): 304-308. |
曾德慧, 尤文忠, 范志平, 等. 2002. 樟子松人工固沙林天然更新障碍因子分析. 应用生态学报, 13(3): 257-261. DOI:10.3321/j.issn:1001-9332.2002.03.001 |
曾东, 李行斌, 于恒. 2000. 新疆落叶松、新疆云杉迹地天然更新特点与规律的辨析. 干旱区研究, 17(3): 46-52. |
Archibold O W, Acton C, Ripley E A. 2000. Effect of site preparation on soil and vegetation cover, and the growth and survival of white spruce (Picea glauca) seedlings, in Saskatchewan. Forest Ecology and Management, 131: 127-141. DOI:10.1016/S0378-1127(99)00205-4 |
Cornett M W, Puettmann K J, Reich P B. 1998. Canopy type, forest floor, predation, and competition influence conifer seedling emergence and early survival in two Minnesota conifer-deciduous forests. Canadian Journal of Forestry Research, 28: 196-205. DOI:10.1139/x97-196 |
Day M E, Schedlbauer J L, Livingston W H, et al. 2005. Influence of seedbed, light environment, and elevated night temperature on growth and carbon allocation in pitch pine (Pinus rigida) and jack pine (Pinus banksiana) seedlings. Forest Ecology and Management, (1/3): 59-97. |
Delong H B, Lieffers V J, Blenis P V. 1997. Microsite effects on first-year establishment and overwinter survival of white spruce in aspen-dominated boreal mixedwoods. Canadian Journal of Forestry Research, 27: 1452-1457. DOI:10.1139/x97-109 |
Facelli J M, Chesson P, Barnes N. 2005. Differences in seed biology of annual plants in arid lands: a key ingredient of the storage effect. Ecology, 86(11): 2998-3006. DOI:10.1890/05-0304 |
Gong Y L, Swqine M D, Miler H G. 1999. Effects of fencing and ground preparation on natural regeneration of native pinewood over 12 years in Glen Tanar, Aberdeenshire. Forestry, 64: 157-168. |
Li W H. 2004. Degradation and restoration of forest ecosystem in China. Forest Ecology and Management, 201: 33-41. DOI:10.1016/j.foreco.2004.06.010 |
Liu Q J. 1997. Structure and dynamics of the subalpine coniferous forest on Changbai mountain, China. Plant Ecology, 132: 97-105. DOI:10.1023/A:1009705319128 |
Liu S R, Li X M, Niu L M. 1998. The degradation of soil fertility in pure larch plantations in the northeastern part of China. Ecological Engineering, 10: 75-86. DOI:10.1016/S0925-8574(97)10024-6 |
McLaren K P, McDonald M A. 2003. The effects of moisture and shade on seed germination and seedling survival in a tropical dry forest in Jamaica. Forest Ecology and Management, 183: 61-75. DOI:10.1016/S0378-1127(03)00100-2 |
Okitsu S, Ito K, Li C. 1995. Establishment processes and regeneration patterns of mountane virgin coniferous forest in northeastern China. Journal of Vegetation Science, 6: 305-308. DOI:10.2307/3236228 |
Schroeder D, Ajith H P. 2002. A comparison of large-scale spatial vegetation patterns following clearcuts and fires in Ontario's boreal forests. Forest Ecology and Management, 159: 217-230. DOI:10.1016/S0378-1127(01)00434-0 |
Sharew H, Legg C J, Grace J. 1997. Effects of ground preparation and microenvironment on germination and natural regeneration of Juniperus procera and Afrocarpus gracilior in Ethiopia. Forest Ecology and Management, 93: 215-225. DOI:10.1016/S0378-1127(96)03962-X |
Tao D L, Xu Z B, Li X. 1987. Effect of litter layer on natural regeneration of companion tree species in the Korean pine forest. Environmental Experimental Botany, 27(1): 53-65. DOI:10.1016/0098-8472(87)90056-6 |
Tsuyuzaki S. 1994. Structure of a thinned Larix olgensis forest in Sandaohu peatland, Jilin Province, China. Natural Areas Journal, 14: 59-60. |
Zhang P C, Shan G F, Zhao G, et al. 2000. China's forest policy for the 21st century. Science, 288(5474): 2135-2136. DOI:10.1126/science.288.5474.2135 |
Zhu J J, Li X F, Liu Z G, et al. 2006. Investigation on the damage of a montane secondary forest induced by snow and wind at Qingyuan experimental forest, Liaoning Province, China. Silva Fennica, 40(1): 37-51. |
Zhu J J, Matsuzaki T, Gonda Y, et al. 2003a. Optical stratification porosity as a measure of vertical canopy structure in a Japanese coastal forest. Forest Ecology and Management, 173(1/3): 89-104. |
Zhu J J, Matsuzaki T, Li F Q, et al. 2003b. Effects of gap size created by thinning on seedling emergency, survival and establishment in a coastal pine forest. Forest Ecology and Management, 182: 339-354. DOI:10.1016/S0378-1127(03)00094-X |