 2007, Vol. 43
2007, Vol. 43文章信息
- 李善文, 张有慧, 张志毅, 安新民, 何承忠, 李百炼.
- Li Shanwen, Zhang Youhui, Zhang Zhiyi, An Xinmin, He Chengzhong, Li Bailian.
- 杨属部分种及杂种的AFLP分析
- AFLP Analysis of Some Species and Hybrids in Populus
- 林业科学, 2007, 43(1): 35-41.
- Scientia Silvae Sinicae, 2007, 43(1): 35-41.
-
文章历史
- 收稿日期:2005-04-01
-
作者相关文章



2. 山东省冠县苗圃 冠县 252500;
3. 美国北卡罗莱纳州立大学林学系 北卡罗莱纳州 NC27695-8203
2. Tree Nursery of Guanxian County, Shandong Province Guanxian 252500;
3. Department of Forestry, North Carolina State University North Carolina State 27695-8203
杨属(Populus)包括白杨派(Leuce)、黑杨派(Aigeiros)、青杨派(Tacamahaca)、胡杨派(Turanga)和大叶杨派(Leucoides),有100余个种,杨属派间、种间存在广泛遗传变异。以往研究杨属派间、种间变异,主要根据形态标记,即杨树的外部形态特征,如树高、胸径、分枝、冠形、叶形、芽形、花形、果形、叶背绒毛、叶缘锯齿等(徐纬英,1988;王明庥等,1991),但是,形态标记数量有限、多态性差、易受环境影响等。随着分子生物学技术的发展,出现了多种分子标记技术,使从基因组水平探讨杨属派间、种间及无性系间遗传变异成为可能(张德强等,2001;张志毅等,2002;Cervera et al., 2005)。与形态标记相比,分子标记具有如下优点:直接以DNA形式表现,在植物的各个组织、器官以及不同发育时期均可检测到,不受季节、环境条件的影响,不存在表达问题;数量多,遍及整个基因组;多态性高;表现中性;有许多分子标记表现为共显性,能够鉴定出纯合基因型与杂合基因型,提供完整的遗传信息等。因此,分子标记被认为是进行遗传变异评价的理想标记(邹喻苹等,2001)。目前,广泛应用的分子标记主要有:RFLP、RAPD、AFLP、SSR等,其中AFLP是一种选择性扩增限制片段的方法,该法集RFLP与RAPD 2种方法的优点于一体,而且能提供比RFLP和RAPD更多的基因组多态性信息(Vos et al., 1995;Mukherjee et al., 2003;Faouzi et al., 2003;Zhang et al., 2004;Chauhan et al., 2004;王献等,2005)。李宽钰等(1996)应用RAPD标记对杨属白杨派、青杨派、黑杨派的20个种的28个无性系研究表明,3个派明显独立,且青杨派与黑杨派关系较近,而与白杨派则相对较远。苏晓华等(1996)对青杨派的甜杨(P. suaveolens)、大青杨(P. ussuriensis)、香杨(P. koreana)和马氏杨(P. maximowiczii)的80个无性系进行RAPD分析,每个树种的全部无性系聚为一类,分子水平分类结果与经典分类一致,大青杨与香杨亲缘关系最近。Michael等(1999)对西班牙的一个胡杨(P. euphratica)群体的257个样品进行AFLP分析,在个体间未检测到任何遗传变异,证明该群体为无性系起源。Maurizio等(2001)应用RAPD标记对53个银白杨(P. alba)家系进行分析并聚类,结果明显分为3类,分别对应于3个不同的地理分布区。
本研究以杨属白杨派、黑杨派、青杨派、胡杨派中的部分种及杂种的47个无性系为试验材料,采用AFLP标记研究它们在DNA水平上的遗传变异,研究杨属无性系间、种间、派间及亲本与子代间亲缘关系;根据分子标记结果探讨杂交亲本间的亲和性和杂交子代早期选择的可行性,讨论分子标记与常规杂交育种的结合点。因此,该项研究具有重要的理论意义和实际应用价值。
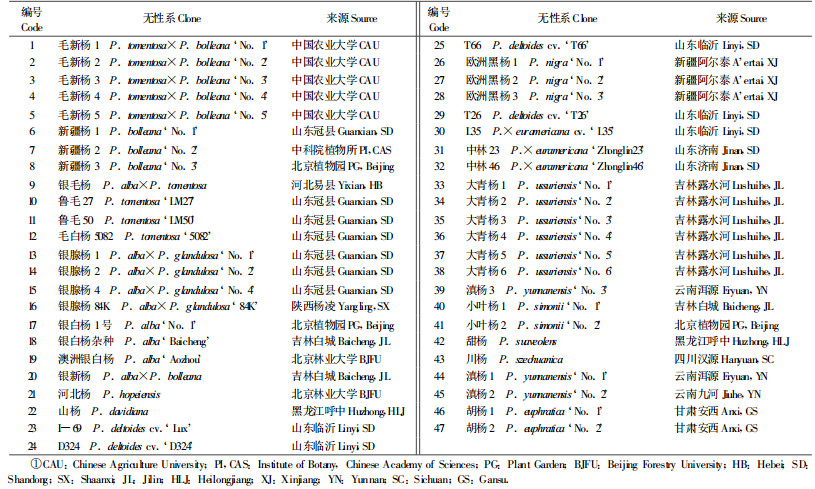
1 材料与方法 1.1 试验材料及其来源试验材料包括杨属中白杨派、黑杨派、青杨派和胡杨派的47个无性系,每个派所包括的无性系及其来源见表 1。
|
|
按Murray等(1980)的方法提取各试验材料的基因组DNA。
1.3 AFLP分析AFLP分析的基本程序和Vos等(1995)发明的方法基本相同,仅对体系进行了优化,采用EcoRⅠ/MseⅠ酶切组合进行基因组限制性酶切,预扩增反应选用引物组合EcoRⅠ+00/MseⅠ+00,选择性扩增反应采用引物组合EcoRⅠ+3/MseⅠ+3。PCR扩增反应在PE-9700PCR仪上进行。筛选的13对引物见表 2。
|
|

选择性扩增产物经变性后在6%的变性聚丙烯酰胺序列分析胶上电泳分离,条件是95 W,3 000 V,恒功率电泳约90 min。电泳后采用Tixier等(1997)的银染检测方法进行AFLP指纹显色反应。Marker为宝生物工程(大连)有限公司生产的100 bp DNA Ladder Marker。
1.5 结果记录统计带型在相同片段位置上的谱带,按0/1系统纪录,有带记为1,无带记为0,并参照Marker带估计片段大小。
1.6 数据统计分析多态性:多态性(%)=扩增多态性片段数/总扩增片段数×100%=(Ni+Nj-2 Nij)/(Ni+Nj-Nij),式中,Nij表示样本i和j的公共带数,Ni、Nj分别是样本i、j的带数。相似系数和遗传距离:采用Nei & Li(1979)的遗传相似系数(genetic similarity,GS),其计算公式为GS=2Nij/(Ni+Nj),式中Nij,Ni,Nj同上;遗传距离D=1-GS。聚类分析:利用NTSYS-pc2.1版分析软件,采用非加权配对算术平均法(Unweighted Pair Group Method Arithmetic Averages,UPGMA)进行聚类。
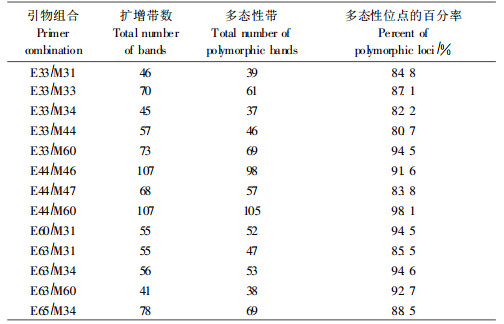
2 结果与分析 2.1 AFLP多态性分析用13对AFLP引物组合对供试样品进行分析,共检测到858个标记,其中多态性标记771个,多态性位点的百分率为89.9%;每对引物组合产生多态性标记数目不等,最少的是E33/M34检测到37条谱带,最多是E44/M60,检测到105条谱带,平均每对引物组合扩增出59个多态性标记,多态性位点的百分率在80.7%~98.1%之间(表 3)。引物组合E44/M46对每个无性系扩增产生的标记位点见图 1。
|
|

|
图 1 AFLP引物组合E44/M46对每个无性系扩增产生的标记位点 Fig. 1 Marker loci generated by AFLP primers combination of E44/M46 for each clone M:100 bp DNA标准100 bp DNA ladder marker. |
根据13对引物扩增结果,计算样本间的遗传距离(李善文,2004),由此数据计算得到各派内样本间、种间遗传距离变幅及其均值(表 4)。
|
|

各样本之间的遗传距离为0.008 0~0.377 4,平均遗传距离为0.172 7;种间遗传距离在0.103 6~0.377 4,均值是0.185 6。种内无性系间遗传距离较小,如毛新杨的5个无性系间的遗传距离在0.019 8~0.064 6,新疆杨的3个无性系间遗传距离变幅为0.008 0~0.015 9;种间遗传距离较大,如山杨与其余样本间的遗传距离在0.271 4~0.377 4。
2.2.2 黑杨派种间和种内无性系间样本间的遗传距离在0.010 3~0.362 1,平均遗传距离为0.192 4;种间遗传距离在0.129 3~0.362 1,均值是0.243 4。种内无性系间遗传距离较小,如美洲黑杨4个无性系间变幅为0.024 9~0.103 1,均值为0.059 3;欧洲黑杨3个无性系间,变幅为0.010 3~0.020 5,均值是0.015 6;3个欧美杨无性系遗传距离在0.027 6~0.076 7,均值是0.056 8;美洲黑杨与欧洲黑杨间遗传距离较大,在0.326 7~0.362 1,均值为0.350 9。
由上述分析又知,美洲黑杨与欧洲黑杨间遗传距离均大于美洲黑杨、欧洲黑杨和欧美杨种内无性系间遗传距离,这一遗传距离(0.350 9)可能与该杂交组合产生杂种优势有密切关系,因为美洲黑杨×欧洲黑杨是世界公认的优良杂交组合,许多著名的欧美杨无性系均来自这一组合,如意大利培育的西玛杨、露易莎杨、Ⅰ-72/58、Ⅰ-45/51、Ⅰ-214等,我国培育的中林46、中林23、中林28、中林14等(李善文等,2004)。
2.2.3 青杨派种间和种内无性系间个体间遗传距离变动于0.028 8~0.386 6,平均遗传距离为0.226 4;种间遗传距离变动于0.130 6~0.386 6,均值为0.248 2。种内无性系间遗传距离较小,如6个大青杨无性系间变幅为0.047 6~0.216 6,均值为0.149 1;3个滇杨无性系的遗传距离在0.028 8~0.064 1,均值为0.047 6。种间遗传距离较大,如川杨与6个大青杨无性系的遗传距离变幅为0.289 3~0.334 5,均值为0.302 1;3个滇杨无性系与6个大青杨无性系的遗传距离变动于0.202 7~0.292 9之间,均值为0.239 7。
2.3 杨属派间遗传变异根据47个样本间的遗传距离计算杨属派间遗传距离(表 5),其中白杨派与胡杨派间最大,为0.531 4;其次是白杨派与黑杨派(0.478 1)、白杨派与青杨派(0.473 2)、黑杨派与胡杨派(0.470 6),这3个遗传距离较接近;再者是青杨派与胡杨派(0.437 9);派间遗传距离最小的是黑杨派与青杨派,为0.326 3。
|
|

用各样本间遗传距离,采用非加权配对算术平均法(UPGMA)进行聚类分析,结果将47个无性系在遗传距离0.140 0处划分为4类(图 2),分别是黑杨派、青杨派、白杨派、胡杨派,这与经典形态分类结果相一致。

|
图 2 杨属47个无性系系统聚类树形图 Fig. 2 System cluster dendrogram of 47 clones in Populus |
在黑杨派内,10个无性系被划分为3类:美洲黑杨、欧美杨和欧洲黑杨。在青杨派中,13个无性系聚类后可分为2类:1)小叶杨、川杨、滇杨类,如果再细分,每个种单独成类;2)大青杨、甜杨类,大青杨6个无性系首先聚在一起,然后与甜杨聚为一类。白杨派内22个无性系聚类后可分为如下3类:1)银白杨、银新杨、银腺杨类;2)毛新杨、新疆杨、毛白杨、河北杨、澳洲银白杨、银毛杨类;3)山杨类。胡杨派的2个无性系聚为一类。
综上分析可知,利用AFLP分子标记遗传距离进行聚类分析,派间聚类结果与形态分类完全一致,派内种间及种内无性系间聚类结果与形态分类也基本相同。这表明应用AFLP分子标记遗传距离对杨属派间、派内种间及种内无性系间的亲缘关系进行定量评价是可行的。
3 结论与讨论利用13对AFLP引物对杨属白杨派、黑杨派、青杨派、胡杨派中19个种的47个无性系进行分析,检测到多态性位点的百分率为89.9%,表明试验材料在分子水平存在广泛变异。聚类分析结果表明,派间聚类与经典形态分类完全一致,派内种间及种内无性系间聚类与形态分类基本相同。李宽钰等(1996)对杨属白杨派、黑杨派、青杨派的20个种28个无性系进行RAPD分析,筛选了7个引物,多态性为79.6%,聚类分析将28个无性系划分3类,分别对应白杨派、黑杨派、青杨派。其试验材料由于未包含胡杨派,因此得到的多态性较低;除选用的试验材料外,不同的分子标记方法也可能是造成多态性差异的原因。
AFLP分析结果与杂交试验、同工酶试验结果基本一致。杨属派间分子遗传距离的大小顺序依次为:白杨派与胡杨派(0.531 4)、白杨派与黑杨派(0.478 1)、白杨派与青杨派(0.473 2)、胡杨派与黑杨派(0.470 6)、胡杨派与青杨派(0.437 9)、黑杨派与青杨派(0.326 3),这说明白杨派与胡杨派、黑杨派、青杨派,胡杨派与黑杨派、青杨派间的亲缘关系较远,这些派间进行杂交不易得到种子,而黑杨派与青杨派间亲缘关系较近,杂交容易成功。该结果与杂交试验和同工酶试验结果相一致,Li等(1981)和Stettler等(1980)的杂交试验表明,白杨派与青杨派、黑杨派间杂交,花粉发育大多不正常,很难得到种子,而黑杨派与青杨派树种杂交则表现出很好的亲和性,且杂种优势明显。徐纬英(1988)以青杨派、黑杨派的种为亲本,从钻天杨×青杨(P. nigra var. italica×P. cathayana)、小叶杨×(钻天杨+旱柳)〔P. simonii×(P. nigra var. italica+Salix matsudana)〕、小叶杨×钻天杨(P. simonii×P. nigra var. italica)组合中分别选出北京杨(P.×beijingensis)、群众杨(P.×popularis)、合作杨(P.×opera)优良品系;王明庥等(1991)以美洲黑杨无性系Ⅰ-69为母本、小叶杨为父本,选育出黑杨派和青杨派间杂种NL-105、NL-106、NL-121等优良无性系。胡志昂(1981)研究了杨属不同派间过氧化物同工酶,结果表明青杨派与黑杨派表现较高的同工酶谱带相似性,而白杨派与青杨派和黑杨派相似性较低。因此,应用AFLP标记分子遗传距离对杨属派间遗传差异进行评价,结果与常规杂交试验和同工酶分析结果相似,说明这种分析方法可行、结果可靠,可以根据亲本间分子遗传距离大小预测杂交亲本间的亲和性。
杂种优良无性系与父母本的遗传距离在不同杂交组合中存在差异。白杨派杂种毛新杨的5个优良无性系与3个新疆杨无性系间的平均遗传距离是0.068 3,与3个毛白杨无性系间的平均遗传距离是0.173 0,这表明毛新杨杂种优良无性系与新疆杨的亲缘关系较近,其相似系数为0.931 7,与毛白杨的亲缘关系较远,相似系数为0.827 0。银毛杨杂种优良无性系与毛白杨3个无性系的遗传距离均值是0.123 9,与银白杨2个无性系的均值是0.126 6,这说明银毛杨杂种优良无性系与母本银白杨、父本毛白杨的亲缘关系相近,其相似系数分别是0.873 4、0.876 1。在黑杨派中,欧美杨3个优良无性系与欧洲黑杨3个无性系的平均遗传距离是0.227 1,与美洲黑杨4个无性系的遗传距离是0.148 4,由此可知,欧美杨杂种优良无性系与美洲黑杨的亲缘关系较近,相似系数为0.851 6,与欧洲黑杨的亲缘关系相对较远,相似系数为0.772 9。本试验所用的毛新杨、银毛杨、欧美杨无性系分别是从毛白杨×新疆杨、银白杨×毛白杨、美洲黑杨×欧洲黑杨的杂种群体中选出的优良无性系,表明有的组合选出的优良无性系与母本相似,有的组合选出的优良无性系与父本相似,有的组合选出的优良无性系属于父母本的中间类型。形态观测结果与这一结论相一致(李善文,2004)。因此,根据亲本和子代的分子遗传距离在不同杂交组合中存在的这种特有关系,可以为其杂交子代相关性状的早期选择提供科学依据。
胡志昂. 1981. 杨属植物的同工过氧化物酶. 植物分类学报, 19(3): 291-296. |
李宽钰, 黄敏仁, 王明庥, 等. 1996. 白杨派、青杨派和黑杨派的DNA多态性及系统进化研究. 南京林业大学学报, 20(1): 6-10. |
李善文, 张志毅, 何承忠, 等. 2004. 中国杨树杂交育种研究进展. 世界林业研究, 17(2): 37-41. DOI:10.3969/j.issn.1001-4241.2004.02.010 |
李善文. 2004.杨树杂交亲本与子代遗传变异及其分子基础研究.北京林业大学博士学位论文 http://cdmd.cnki.com.cn/Article/CDMD-10022-2005076735.htm
|
苏晓华, 张绮纹, 张望东, 等. 1996. 大青杨及其近缘种的遗传变异和系统关系研究. 林业科学, 32(2): 118-124. |
王明庥, 黄敏仁, 邬荣领, 等. 1991.美洲黑杨×小叶杨杂交育种研究//林业部科技司.阔叶树遗传改良.北京: 科学技术文献出版社, 83-92
|
王献, 张启翔, 杨秋生, 等. 2005. 利用AFLP研究紫薇的亲缘关系. 北京林业大学学报, 27(1): 59-63. DOI:10.3321/j.issn:1000-1522.2005.01.013 |
徐纬英. 1988. 杨树. 哈尔滨: 黑龙江人民出版社.
|
张德强, 张志毅, 杨凯. 2001. 分子标记技术在杨树遗传变异及系统分类中的应用. 北京林业大学学报, 23(1): 76-80. DOI:10.3321/j.issn:1000-1522.2001.01.019 |
张志毅, 林善枝, 张德强, 等. 2002. 现代分子生物学技术在林木遗传改良中的应用. 北京林业大学学报, 24(5/6): 250-261. |
邹喻苹, 葛颂, 王晓东. 2001. 系统与进化植物学中的分子标记. 北京: 科学出版社.
|
Cervera M T, Storme V, Soto A, et al. 2005. Intraspecific and interspecific genetic and phylogenetic relationships in the genus Populus based on AFLP markers. Theor Appl Genet, 111: 1440-1456. DOI:10.1007/s00122-005-0076-2 |
Chauhan N, Negi M S, Sabharwal V, et al. 2004. Screening interspecific hybrids of Populus (P.ciliata×maximowiczii) using AFLP markers. . Theor Appl Genet, 108: 951-957. DOI:10.1007/s00122-003-1508-5 |
Faouzi B, Bruce M, Bill S. 2003. Application of DNA markers for the identification and management of hybrid poplar accessions. Agroforestry System, 59: 53-59. DOI:10.1023/A:1026189103893 |
Li Wendian, Xu Han. 1981. The behavior of pollen tubes on the stigma in the intersectional crosses in Populus and its relation to seed-setting. Acta Botanica Sinica, 3(2): 102-109. |
Maurizio S, Renato D O, Oronzo A T, et al. 2001. Assessment of geographic variation by RAPD markers among Italian open-pollinated progenies of Populus alba L. Genetic Resources and Crop Evolution: 1-6. |
Michael F F, Dolores M L, Molly M K, et al. 1999. From the waters of Babylon? Populus euphratica in Spain is clonal and probably introduced. Biodiversity and Conservation: 769-778. |
Mukherjee A K, Acharya L K, Mattagajasingh I, et al. 2003. Molecular characterization of three Heritiera species using AFLP markers. Biologia Plantarum, 47(3): 445-448. |
Murray M G, Thompson W F. 1980. Rapid isolation of high-molecular-weight plant DNA. Nucleic Acids Res, 8: 4321-4325. DOI:10.1093/nar/8.19.4321 |
Nei M, Li W H. 1979. Mathematical model for studying genetic variation in term of restriction endonucleases. Proc Natl Acad Sci, 76: 5269-5273. DOI:10.1073/pnas.76.10.5269 |
Stettler R F, Kostob R, Steenockers V. 1980. Interspecific crossability studies in poplars. Theor Appl Genet, 58: 273-282. DOI:10.1007/BF00265179 |
Tixier M H, Sourdille R M, Leroy P, et al. 1997. Detection of wheat microsatellites using a non-radioactive silver-nitrate staining method. J Genet Breed, 51: 175-177. |
Vos P, Hogers R, Bleeker M, et al. 1995. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res, 23: 4407-4414. DOI:10.1093/nar/23.21.4407 |
Zhang Deqiang, Zhang Zhiyi, Yang Kai, et al. 2004. Genetic mapping in (Populus tomentosa×Populus bolleana) and P.tomentosa Carr. using AFLP markers. Theor Appl Genet, 108: 657-662. DOI:10.1007/s00122-003-1478-7 |