 2006, Vol. 42
2006, Vol. 42文章信息
- 喻方圆, 邵岚, 沈永宝.
- Yu Fangyuan, Shao Lan, Shen Yongbao.
- 杉木、马尾松种子贮藏过程中生理生化变化研究
- Studies on Physiological and Biochemical Changes of Chinese Fir and Masson Pine Seeds during Storage
- 林业科学, 2006, 42(12): 137-142.
- Scientia Silvae Sinicae, 2006, 42(12): 137-142.
-
文章历史
- 收稿日期:2005-11-08
-
作者相关文章


杉木(Cunninghamia lanceolata)、马尾松(Pinus massoniana)是我国南方主要造林树种,每年造林用种量很大。由于杉木、马尾松结实都存在大小年现象,大年结实量多,播种品质好,采种成本低;小年结实量少,播种品质差,采种成本高。因此,生产上常常贮藏杉木、马尾松种子,以达到以丰补歉的目的。近年来,随着生物遗传多样性的日益被重视,作为种质资源保存措施之一,杉木、马尾松种子的长期贮藏也被提上议事日程。因此,研究杉木、马尾松种子的贮藏问题具有十分重要的意义。
尽管对杉木、马尾松种子贮藏问题的研究有不少报道,但这些研究基本上只能解决种子短期贮藏的问题,研究方法比较简单,对种子耐藏性机理的研究不够深入。如果从生理生化的角度,对杉木、马尾松种子耐藏性的机理进行深入研究,从而揭示杉木、马尾松种子耐藏性的机理,并在此基础上制定杉木、马尾松种子贮藏的策略,则无论对商品种子生活力的保持,还是对种质资源的长期保存,都具有重要的理论和实际意义。
本文旨在通过对不同贮藏条件下种子发芽率、细胞膜透性、抗氧化系统酶活性等生理生化指标的测定,研究贮藏条件对杉木、马尾松种子生理生化特性的影响,为杉木、马尾松种子贮藏提供理论依据。
1 材料与方法 1.1 试验材料杉木种子2002年采自江西省吉安县,马尾松种子2002年采自广西壮族自治区藤县。种子样品基本情况见表 1。
|
|
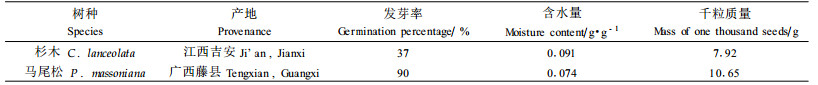
1) 种子贮藏条件的设置 杉木种子的含水量为3种处理,即C1、C2和C3,分别为0.038 g·g-1(4%)、0.091 g·g-1(10%)和0.153 g·g-1(18%);马尾松种子的含水量也为3种处理,即P1、P2和P3,分别为0.038 g·g-1(4%)、0.074 g·g-1(8%)和0.123 g·g-1(14%),将他们分别贮藏在25 ℃、3 ℃、-18 ℃和-30 ℃条件下,采用两层聚乙烯塑料袋密封包装, 贮藏期间保持种子含水量基本不发生变化。每隔120 d取样1次,用于测定生理生化指标。2)种子发芽率的测定 采用《林木种子检验规程》(国家质量技术监督局,2000)中的方法测定种子发芽率。3)种子浸出液相对电导率的测定 分别称取不同贮藏条件下的种子1 g,先用清水冲洗3次,再用蒸馏水冲洗2次,然后放入100 mL的三角瓶中,加入50 mL蒸馏水,25 ℃恒温浸泡12 h,用DDS-ⅡA型电导仪测定电导值(S1)。测毕,用塑料薄膜封瓶口,将浸泡液连同种子在沸水中煮30 min,冷却至室温测定其电导值(S2)。以不加种子的重蒸馏水为空白对照。按下式计算出相对电导率(L)。
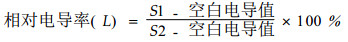
|
4) 抗氧化系统酶活性的测定 取不同处理的种子1 g,吸胀12 h,加5 mL K2HPO4-KH2PO4缓冲液(50 mmol·L-1,pH 7.0)冰浴,研磨成匀浆。在4 ℃的条件下,15 000 g离心20 min。上清液即为酶提取液。采用王爱国等(1983)的氯化硝基氮蓝四唑(NBT)光化学还原法测定超氧化物歧化酶(SOD)活性;采用愈创木酚法(李合生等,2000)测定过氧化物酶(POD)活性。5)丙二醛(MDA)含量的测定 采用TBA(硫代巴比妥酸)法测定丙二醛(李合生等,2000)含量。
1.3 数据分析采用SAS6.12和Excel软件对实验数据进行统计分析。
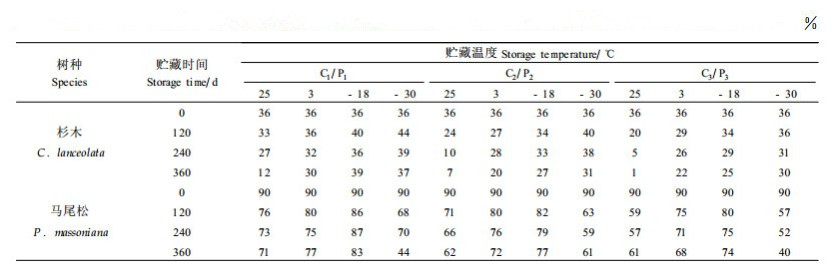
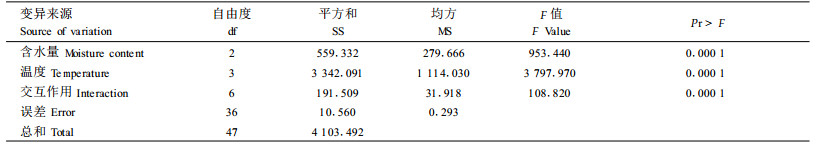
2 结果与分析 2.1 不同贮藏条件对杉木、马尾松种子发芽率的影响本试验测定了不同含水量和贮藏温度条件下,杉木、马尾松种子发芽率的变化情况,结果见表 2。方差分析的结果表明,不同的温度和含水量对杉木、马尾松种子发芽率的影响非常明显,差异达到显著水平(表 3、4)。
|
|
|
|
|
|
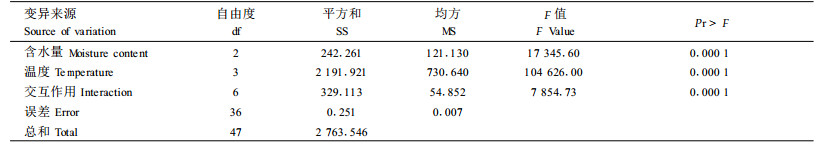
从表 2中可以看出,在种子含水量相同的情况下,贮藏温度对杉木种子发芽率有显著影响, 贮藏温度越高,种子发芽率越低。在温度相同的情况下,不同含水量对种子发芽率也有显著影响。值得注意的是,贮藏在25 ℃条件下的杉木种子,无论含水量如何,贮藏360 d后,其发芽率的下降均非常明显,但不同含水量种子发芽率大幅下降的起点不同,其中含水量18%的种子从贮藏开始起,发芽率就大幅下降,含水量10%的种子发芽率大幅下降的起点是贮藏120 d以后,而含水量4%的种子发芽率大幅下降的起点是贮藏240 d以后。这一结果突出说明了温度和含水量对种子贮藏寿命的主导作用。
从马尾松种子的情况来看,贮藏在-18 ℃条件下种子发芽率下降最为平缓,其次3 ℃,再次25 ℃,-30 ℃下降最为明显。含水量4%的马尾松种子在-30、-18、3、25 ℃条件下贮藏3 60 d后,发芽率分别比贮藏前下降了46%、7%、13%、16%。马尾松种子含水量为8%和14 %时,温度对其发芽率的影响与在4%含水量条件下趋势相似,贮藏360 d后,-18 ℃条件下贮藏的含水量8%和14%的马尾松种子发芽率分别是贮藏在25 ℃下的1.24倍和1.21倍,是贮藏在-30 ℃下的1.26倍和1.85倍。值得注意的是马尾松种子在-30 ℃的条件下贮藏效果最差,且在贮藏初期,发芽率就有较大幅度的下降,可能的原因是马尾松种子在-30 ℃的条件下遭受了冻害。
与杉木种子相比,无论何种温度与含水量的组合,贮藏360 d后,马尾松种子的发芽率都没有出现大幅度下降的现象,说明马尾松种子的耐藏性优于杉木种子。
2.2 不同贮藏条件对杉木、马尾松种子浸出液相对电导率的影响本文测定了贮藏在不同的温度和含水量条件下杉木、马尾松种子浸出液相对电导率的变化情况,结果见表 5。从表 5中可以看出,随着贮藏时间的延长,种子浸出液的相对电导率都呈现出逐渐升高的趋势,在含水量相同的条件下,不同贮藏温度种子浸出液相对电导率上升的趋势有所不同。温度和含水量对贮藏360 d后杉木、马尾松种子浸出液相对电导率影响的方差分析结果表明,不同温度和含水量对杉木、马尾松种子浸出液相对电导率的影响均有显著差异(表略去)。
|
|
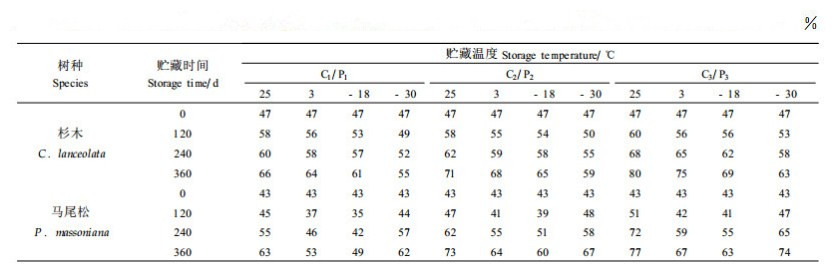
杉木种子浸出液相对电导率随贮藏时间的延长而增加的趋势总体比较平缓,不同贮藏温度下种子浸出液相对电导率上升的幅度不同,25 ℃条件下上升的幅度最大,3 ℃其次,-18、-3 0 ℃依次减缓。不同含水量种子浸出液相对电导率随贮藏时间的延长而增加的趋势也有差异,含水量为4%的条件下,种子浸出液相对电导率增加的速率最小,含水量10%的种子次之,含水量18%的种子浸出液相对电导率增加的速率最大。
从马尾松种子的情况来看,不同贮藏条件对种子浸出液相对电导率的影响与杉木种子有所不同,在贮藏的前120 d时间内,种子浸出液相对电导率变化平缓,甚至有所下降。但随贮藏时间的延长,种子浸出液相对电导率不断上升,其中不同贮藏温度上升的幅度有显著差异。值得注意的是,-30 ℃条件下种子浸出液相对电导率与25 ℃条件下趋势接近,说明在这两种温度条件下种子细胞膜随贮藏时间的延长都受到了较大的损害。在种子含水量方面,随贮藏时间的延长,种子含水量越高,种子细胞浸出液相对电导率上升的幅度越大。
2.3 不同贮藏条件对杉木、马尾松种子SOD活性的影响本文测定了贮藏在不同的温度和含水量条件下,杉木、马尾松种子SOD活性的变化情况,结果见表 6。从表 6中可以看出,种子在不同条件下贮藏360 d的过程中,随着贮藏时间的延长S OD酶活性逐渐降低。方差分析结果(表略)表明,不同温度和含水量对杉木、马尾松种子SOD酶活性影响非常明显,差异达到显著水平。
|
|
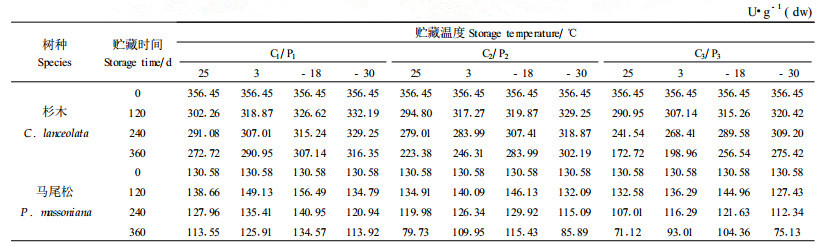
从表 6中可以看出,当含水量为18%的杉木种子贮藏120 d后,不同温度下SOD活性之间的差异便明显地表现出来,贮藏360 d后的种子在25、3、-18和-30 ℃条件下的SOD活性分别比贮藏前降低了2.58倍、1.97倍、1.45倍和1.29倍。含水量10%的种子贮藏240 d后,各温度之间酶活性明显表现出差异,而含水量4%的种子在贮藏360 d的时间内差异不是很显著。说明种子含水量在杉木种子贮藏过程中对SOD活性的变化起重要作用。从表 6中还可以看出,马尾松种子SOD活性在贮藏开始的120 d内几乎没有变化,之后酶活性才缓慢下降。其中不同含水量的种子下降幅度不同,含水量14%的种子下降幅度最大,含水量10%的种子次之,含水量4%的种子下降最小。从贮藏温度对马尾松种子SOD活性的影响来看,含水量4%的种子差异不明显,含水量10%和14%的种子在贮藏过程中不同温度间的差异逐渐显现出来,其中-30 ℃条件下贮藏的种子SOD活性下降最多,25 ℃次之,-3和-18 ℃条件下贮藏的种子下降幅度较小。
2.4 不同贮藏条件对杉木、马尾松种子POD活性的影响本文测定了贮藏在不同的温度和含水量条件下杉木、马尾松种子POD活性的变化,结果见表 7。方差分析结果(表略)表明,不同的温度和含水量对杉木、马尾松种子POD酶活性的影响非常明显,差异达到显著水平。
|
|
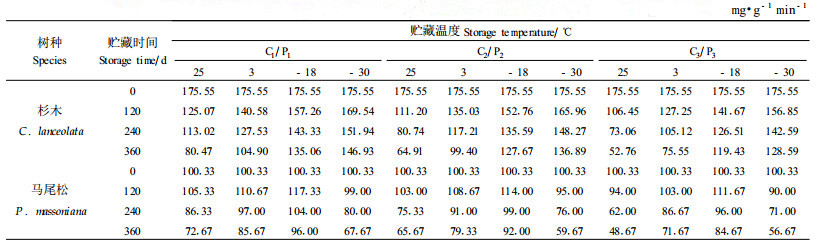
从表 7中可以看出,不同贮藏温度条件下,杉木种子的POD活性差异较为明显,贮藏温度越高,相同含水量种子的POD活性下降低速度越快,-30 ℃条件下POD活性下降幅度最小,-18 ℃次之,3 ℃和25 ℃条件下POD活性下降较多。从表 7中还可以看出,马尾松种子在贮藏120 d的时间内POD活性变化不大,贮藏120 d以后开始下降,且贮藏温度不同,下降的幅度不同。其中贮藏在-18 ℃和3 ℃条件下的马尾松种子SOD活性下降较少,25 ℃和-30 ℃条件下的种子POD活性下降较多。
2.5 不同贮藏条件对杉木、马尾种子丙二醛含量的影响本文研究了贮藏在不同的温度和含水量条件下杉木、马尾松种子MDA含量的变化,结果见表 8。方差分析结果(表略)表明,不同的温度和含水量对杉木、马尾松种子的丙二醛含量的影响非常明显,差异达到显著水平。
|
|
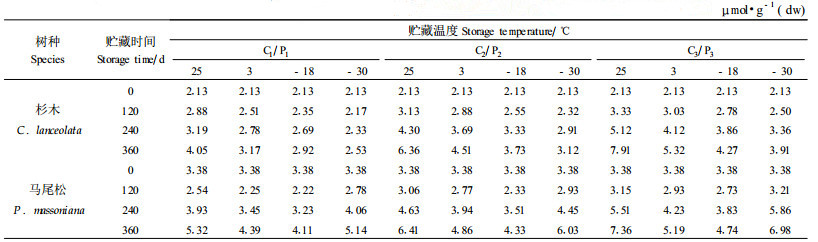
从表 8中可以看出,不同含水量条件下杉木种子MDA含量随贮藏时间的延长均有上升的趋势,但较为平缓。只有贮藏在25 ℃条件下的杉木种子,无论含水量如何,其MDA含量随贮藏时间的延长均有较强的上升趋势。当然,含水量越高,上升趋势越强。当贮藏温度为25 ℃时,种子贮藏360 d后,含水量18%、10%和4%的杉木种子MDA含量分别比贮藏前提高了3.91倍、2.89倍和1.70倍。
从表 8中还可以看出,不同含水量条件下的马尾松种子在不同贮藏温度下MDA含量随贮藏时间的延长而变化的趋势均为先下降再上升,MDA含量在贮藏120 d后达到最小值,以后不断上升。其中不同贮藏温度条件下上升的幅度不同25 ℃和-30 ℃条件下上升较多,3 ℃和-18 ℃条件下上升较少。不同含水量对马尾松种子MDA含量也有显著影响,含水量4%的种子MDA含量最小,其次为含水量10%的种子,含水量14%的种子MDA含量最高。
3 讨论1) 种子含水量和贮藏温度是影响种子贮藏寿命的关键因素(Harrington,1972)。Ellis等(1980;1986)的研究证明,通过降低种子含水量,某些正常型种子可以在适当提高贮藏温度的条件下,达到较高含水量和低温条件下相近的贮藏效果。在此基础上,Bonner(1999)建立了火炬松、湿地松、北美枫香和美国悬铃木(Plantanus occidentalis)等林木种子的贮藏寿命预测方程。本文通过研究不同含水量杉木、马尾松种子在不同贮藏温度条件下种子发芽率的变化,得到的结果与Ellis等(1980;1986)的研究有相同的趋势,即种子的含水量和贮藏温度越低,发芽率下降的速度越缓慢。但要建立杉木、马尾松种子贮藏寿命的预测方程,还需要贮藏更长时间的试验研究。本研究中值得注意的是,马尾松种子贮藏在-30 ℃的低温条件时,出现了种子发芽率大幅下降的现象,可能的原因是马尾松种子细胞膜脂在-30 ℃的低温条件产生相变,导致细胞膜受到损伤。这说明Ellis等(1980;1986)的种子活力方程在应用时是有一定限度的,如对马尾松种子而言,在-30 ℃的低温条件下,并不是贮藏温度越低,种子的贮藏寿命越长。
2) Mcdonald(1999)指出,脂质过氧化是种子劣变的主要原因之一。在种子贮藏过程中,膜脂不饱和脂肪酸过氧化,使得原来膜上排列整齐的磷脂分子变得疏松且不整齐,导致细胞膜完整性遭破坏,在一定条件下贮藏超过一定期限后,膜的结构和功能就会丧失,导致种子活力降低以至丧失。本文通过对不同含水量和贮藏温度条件下杉木、马尾松种子浸出液相对电导率和MDA含量的研究,证实种子贮藏过程中膜脂确实发生了过氧化作用。值得注意的是,杉木种子膜脂发生劣变的时间早于马尾松种子,说明杉木种子的耐藏性不如马尾松种子。在本文对种子发芽率的研究中发现,无论何种温度与含水量的组合,贮藏360 d后,马尾松种子的发芽率都没有出现大幅度下降的现象,正好印证了杉木种子膜脂发生劣变的时间早于马尾松种子这一现象。
3) SOD和POD是种子老化过程中清除自由基的关键酶类,对延长种子贮藏寿命具有重要的意义。但种子老化过程中SOD和POD的含量是上升还是下降,不同研究者有不同的说法。如Kalpana等(1994)研究加速老化木豆种子SOD的变化后认为,在加速老化的过程中,木豆种子的SOD含量上升。而Bailly等(1997)对向日葵的研究却认为,种子加速老化过程中SO D和CAT的含量下降。本文的研究认为,杉木、马尾松种子老化过程中的SOD和POD含量是不断下降的,SOD和POD含量的变化与种子贮藏寿命有密切关系。因种子内保护酶系统清除自由基的能力不足,致使自由基产生与清除的平衡被打破,自由基不断积累攻击膜脂分子,引起过氧化作用,最终导致种子劣变。这可能是种子老化的重要内部原因。
4) 王培蒂等(1994)对马尾松种子进行长期贮藏研究后发现,种子贮藏初期由于种子的后熟发芽率有所升高,而且产地所处的气候带不同,其后熟年限有所差别。本研究也发现,在25、3和-18 ℃的条件下贮藏120 d的马尾松种子,其发芽率、SOD和POD活性均有不同程度的提高,说明马尾松种子确实存在后熟现象。
5) 在本文的研究中,发现贮藏在不同含水量和温度条件下的种子生理生化指标之间有些差异不显著,比如,含水量4%的杉木种子在-18 ℃和-30 ℃之间差异不显著,含水量4%的马尾松种子在3 ℃和-18 ℃之间差异不显著。这是因为种子老化是一个渐进、缓慢的过程,特别是在温度和含水量均较低的贮藏条件下。对于杉木、马尾松种子而言,经过360 d的贮藏,其贮藏寿命的差异并没有完全显现,但相信随着贮藏时间的延长,他们之间的差异会变的更大。
国家质量技术监督局. 2000. 中华人民共和国国家标准《林木种子检验规程》(GB2772-1999)北京: 中国标准出版社, 10-15 http://www.csres.com/detail/61893.html
|
李合生, 孙群, 赵世杰, 等. 2000. 植物生理生化实验原理和技术. 北京: 高等教育出版社, 105-261.
|
王爱国, 罗广华, 邵从本, 等. 1983. 大豆种子超氧物歧化酶的研究. 植物生理学报, 9(1): 77-83. |
王培蒂, 秦国峰. 1994. 长期贮藏对马尾松种子品质的影响. 林业科学研究, 7(2): 193-198. |
Bailly C, Benamar A, Corbineau F, et al. 1997. Changes in superoxide dismutase, catalase, and glutathione reductase activities in sunflower seeds during accelerated aging and subsequent priming. In: Basic and Applied Aspects of Seed Biology (eds. Ellis R H, Black M, Murdoch A J, et al), Boston: Kluwer Academic Publisher. 349-360
|
Bonner F T. 1999. Viability equations for forest tree seeds. Seed Science and Technology, 27: 981-989. |
Ellis R H, Hong T D, Roberts E H. 1986. Logarithmic relationship between moisturecontent and longevity in sesame seeds. Annals of Botany, 57: 499-503. DOI:10.1093/oxfordjournals.aob.a087131 |
Ellis R H, Roberts E H. 1980. Improved equations for the prediction of seed longevity. Annals of Botany, 45: 13-30. DOI:10.1093/oxfordjournals.aob.a085797 |
Harrington J F. 1972. Seed storage and longevity. In: Kozlowski (ed) Seed Biology. New York and London, (3): 145-245
|
Kalpana R, and Madhava Rao K V. 1994. Absence of the role of lipid peroxidation during accelerated aging of seeds of pigeonpea (Cajanus cajan (L.) Millsp.) cultivars. Seed Science and Technology, 22: 253-260. |
McDonald M B. 1999. Seed deterioration: physiology, repair and assessment. Seed Science and Technology, 27: 177-237. |