 2006, Vol. 42
2006, Vol. 42文章信息
- 何友均, 崔国发, 邹大林, 郑杰, 董建生, 李永波, 郝万成, 李长明.
- He Youjun, Cui Guofa, Zou Dalin, Zheng Jie, Dong Jiansheng, Li Yongbo, Hao Wancheng, Li Changming.
- 三江源自然保护区玛珂河林区寒温性针叶林优势灌木种间联结研究
- Interspecific Association of Dominant Shrub Species in Cold Temperate Coniferous Forest in Ma Kehe Region of San Jiangyuan National Nature Reserve
- 林业科学, 2006, 42(12): 126-129.
- Scientia Silvae Sinicae, 2006, 42(12): 126-129.
-
文章历史
- 收稿日期:2005-11-07
-
作者相关文章



2. 中国林业科学研究院林业科技信息研究所 北京 100091;
3. 北京市怀柔区林业局 北京 101400;
4. 青海省林业局 西宁 810007;
5. 青海省玛珂河林业局 班玛 814300
2. Research Institute of Forestry Policy and Information, Chinese Academy of Forestry Beijing 100091;
3. Forestry Bureau of Huairou District in Beijing Beijing 101400;
4. Forestry Bureau of Qinghai Province Xining 810007;
5. Ma Kehe Forestry Bureau, Qinghai Province Banma 814300
种间联结是不同物种在空间分布上的相互关联性,通常是由群落生境的差异影响了物种的分布而引起的(Greig-Smith,1983)。种间联结研究在理论上有助于了解物种间的相互关系,正确认识群落的结构特征,探讨环境差异对植物群落的影响,提供了一种客观认识自然种群的方法,为物种多样性保护提供了科学依据(蒋有绪,1982;史作民,1998;彭少麟等,1999)。
玛珂河林区是三江源国家级自然保护区的核心区之一,林区中的森林植被类型主要是寒温性针叶林,主要包括川西云杉(Picea balfouriana)林、紫果云杉(Picea purpurea)林、鳞皮云杉(Picea retroflexa)林、大果圆柏(Sabina tibetica)林、密枝圆柏林(Sabina convallium)、鳞皮冷杉(Abies faxoniana)林等。寒温性针叶林的林相整齐,树木分布较均匀,大环境相似;寒温性针叶林中灌木种类较多,尤其是优势灌木对寒温性针叶林的演替和恢复有着十分重要的影响。为了弄清该林区优势灌木物种间的相互关系及其在利用资源和占据生态位中的功能和地位,本文采用种间联结相关技术,对寒温性针叶林中优势灌木种群的种间联结关系进行了分析。
1 材料与方法 1.1 研究区自然概况青海省玛珂河林区位于果洛藏族自治州班玛县境内(32°15′—32°30′N,99°53′—100°31′E)。最高海拔5 300 m,最低海拔3 147 m。森林土壤属于暗棕色针叶林土。属凉温半湿润气候类型,年均气温0.1~2.5 ℃,7月平均气温9.6~12.4 ℃,1月平均气温-11.2~-7.0 ℃,全年极端最高气温28.1 ℃,全年极端最低气温-29.7 ℃;年降水量652.4~764.4 mm,70%以上集中在6—9月份。主要植被类型为寒温性针叶林、高寒灌丛和高寒草甸。
1.2 野外调查与取样尺度对植物群落种间关系的格局具有重要的决定性作用,在较大尺度内,环境的异质性对种间关联性起主要作用,而在较小尺度内,种间竞争则起主导作用(Jonsson et al., 1998)。一般认为,取样面积过大,则可能多是正联结,而取样面积过小,则又可能形成负联结过多的情况(刘金福,2001)。因此,取样时应考虑研究区域内物种和生境的变化幅度来确定群落的最小适合面积。
根据三江源自然保护区玛珂河林区所处青藏高原独特的地理位置和环境对植物生长的影响,结合种-面积曲线取样原理,2003年7—9月,在三江源自然保护区玛珂河林区采用典型抽样和标准地调查法,在寒温性针叶林中共设置20 m×20 m的样地40块,然后在每一块样地的四角和中央共设置5个2 m×2 m的灌木样方。对样地内的乔木进行每木检尺,测定其胸径、树高、株数;测定和记录样方内灌木的种类、株数、高度和盖度。同时,记录样地所处的海拔、坡度和坡向等生态因子指标。
1.3 数据处理在40块样地中选取20种优势灌木作为种间联结的研究对象,植物物种名称和代码见表 1。根据样地调查资料,统计分析各灌木物种共存与否的样方数,组成40×20灌木多度数据矩阵,当第i物种在第j样方出现时,记为1,否则记为0,将定量多度数据矩阵转化为二元数据(0,1)矩阵,以此作为种间联结分析的原始数据。
|
|
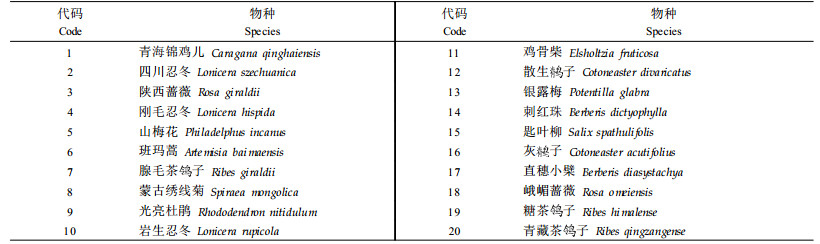
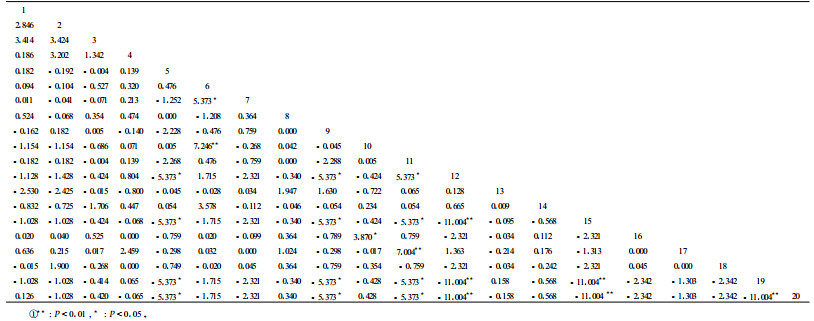
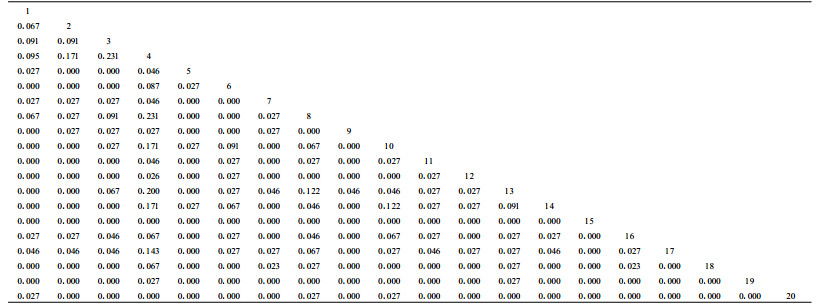
χ2统计量检测公式为χ2={[(ad-bc)-0.5N]2N}/{(a+b)(a+c)(b+d)(c+d)}。式中:N为取样总数;a表示物种A和B同时出现;b表示物种A出现而B不出现;c表示物种A不出现而物种B出现;d表示物种A和B都不出现。由于关联有两种类型,若(ad-bc)>0为正联结,若(ad-bc)<0为负联结,因此χ2>χ0.052(1)为种对联结性显著,否则不显著。χ0.052(1)=3.841,χ0.012(1)=6.635,若3.841<χ2<6.635为联结显著;若χ2>6.635为极显著,否则不显著。优势灌木χ2统计量结果见表 2。共同出现百分率PC计算公式为PC=a/(a+b+c)。PC的值域为[0,1]。其值越趋近于1,表明该物种对的正联结越紧密(王伯荪等,1985;彭少麟等,1999)。优势灌木共同出现百分率PC见表 3。
|
|
|
|
从表 2可知,20种灌木,共有190对种间组合,其中有2对极显著正联结(P<0.01),3对显著正联结(P<0.05)。说明这些物种常常相伴出现,主要是由于这些物种具有相似的生态学特性,对环境有相似的要求,都选择或避免相同的生态因子,在一定程度上体现了生态位的重叠性(马丹炜,2004)。例如6(班玛蒿)与10(岩生忍冬)呈极显著正联结,主要因为2者对生境有相同的生态适应性,班玛蒿与岩生忍冬经常出现在川西云杉林中;11(鸡骨柴)与17(直穗小檗)也呈极显著正联结,能在水分条件较差的阳坡地段生长,对生境条件要求相似,虽然在资源利用上可能存在竞争,但在养分和空间资源充足的情况下,就不会相互排斥,并且同时出现的机率非常大,呈极显著正联结。6(班玛蒿)与7(腺毛茶鸰子)呈显著正联结,共同出现在川西云杉天然林内,其中班玛蒿是林内灌木层的优势种,而腺毛茶鸰子的高度大于班玛蒿高度,但数量较少,他们能够同时利用不同空间的生态位而呈正联结;10(岩生忍冬)与16(灰鹡子)呈显著正联结,通过分析测试,发现他们在川西云杉林和鳞皮云杉林中共同出现的机率最大,而且是这两种林分灌木层的优势种,比较喜欢阴凉的生境,能够共同利用有限的资源;11(鸡骨柴)与12(散生鹡子)都出现在密枝圆柏林中,成为灌木层的共优种,都喜欢生长在阳光充足的地段,由于密枝圆柏林盖度较小,外界的阳光能够直射林中,因此鸡骨柴和散生鹡子都能够利用充足的阳光,种间竞争不激烈,能够共同生长在同一林分中而表现出正联结。
表 2表明,有6对灌木物种呈极显著负联结(P<0.01),11对呈显著负联结(P<0.05),其共同出现百分率均为0(表 3),这是他们长期适应微环境而对生境需求不一致或具有不同的生物学和生态学特性的结果,也是他们利用不同资源空间和对有限资源竞争的结果,同时还是生态位分离的反映(李先琨等,1999),因而表现出显著和极显著的负联结。例如9(光亮杜鹃)与12(散生鹡子)、9(光亮杜鹃)与15(匙叶柳)、9(光亮杜鹃)与19(糖茶鸰子)、9(光亮杜鹃)与20(青藏茶鸰子)之间表现出显著负联结,以及12(散生鹡子)与15(匙叶柳)、12(散生鹡子)与19(糖茶鸰子)、12(散生鹡子)与20(青藏茶鸰子)相互之间表现出极显著负联结,主要是这些物种生态位重叠机会很小,甚至没有重叠,各自占据一定的生态位。具体而言,光亮杜鹃和散生鹡子都分布在密枝圆柏林中,都比较喜欢阳光充足的地方,而且都能在密枝圆柏林中生存,对生境要求有一定的相似性,他们表现出显著负联结的原因可能是对有限资源强烈竞争的结果;匙叶柳生长在较为干旱的地段,糖茶鸰子和青藏茶鸰子都生长在水分条件较好的寒温性针叶林中,可见光亮杜鹃和散生鹡子分别与匙叶柳、糖茶鸰子和青藏茶鸰子的显著性负联结,主要原因是对水分条件的要求不同;虽然糖茶鸰子和青藏茶鸰子都生长在水分较好的生境中,但他们竞争有限的资源而相互排斥,表现为极显著负联结。
从表 2还可以看出,完全独立的有8对,占总数的4.21%,说明这些灌木物种之间对生态环境的需求不同,虽然有些物种生态学特性和生物学特性相似,但是为了生存,相互之间竞争有限的资源而独立占据一定生态位。
表 3是寒温性针叶林优势灌木种间共同出现百分率半矩阵。共同出现百分率与种间联结系数都能用来测定物种间的联结性的大小。共同出现百分率的值愈高,物种同时出现的机率愈大,但并非一定指示正联结;反之,其值低,只说明物种之间同时出现的机率低,但并非一定意味着负联结。从表 3可以看出,大部分物种之间共同出现百分率均为0,且其他物种间共同出现百分率也较小,进一步说明该林区寒温性针叶林优势灌木物种之间共同出现的机率较小。
3 结论取样过程中综合考虑青藏高原物种和生境的变化幅度,结合种-面积曲线取样原理,确定样地最小适合面积为20 m×20 m,样地内灌木样方大小为2 m×2 m,从优势种间关系研究结果来看,与三江源自然保护区玛珂河林区所处青藏高原独特的地理位置和区域气候形成的植物种间关系相一致。
寒温性针叶林中的190对优势灌木组合,仅有69对呈现正联结,占总数的36.32%,其中有2对呈极显著正联结(P<0.01),3对呈显著正联结(P<0.05)。负联结有113对,占总数的59.47%,其中有6对灌木物种呈极显著负联结(P<0.01),11对呈显著负联结(P<0.05),他们的共同出现百分率均为0;完全独立的有8对,占总数的4.21%。研究表明,研究区域的多数灌木物种对生态环境的需求不同,虽然有些物种的生态学特性和生物学特性相似,但是为了生存,相互之间竞争有限的环境资源和空间资源而产生竞争和排斥,表现为生态位分离;仅有少数优势灌木物种对环境有相似的要求,能够共同利用有效资源,在一定程度上体现了生态位的重叠性。
蒋有绪. 1982. 川西亚高山森林植被的区系、种间关联和群落排序的生态分析. 植物生态学与地植物学丛刊, 6(4): 281-300. |
李先琨, 黄玉清, 苏宗明. 1999. 南方红豆杉群落主要树木种群间联结关系初步研究. 生态学杂志, 18(3): 10-14. DOI:10.3321/j.issn:1000-4890.1999.03.003 |
刘金福. 2001. 天然格氏栲林乔木层种群种间关联性研究. 林业科学, 37(4): 117-123. DOI:10.3321/j.issn:1001-7488.2001.04.019 |
马丹炜. 2004. 青城山森林植被常见种群种间联结性的研究. 四川大学学报:自然科学版, 41(1): 169-173. |
彭少麟, 周厚诚, 郭少聪, 等. 1999. 鼎湖山地带性植被种间联结变化研究. 植物学报, 41(11): 1239-1244. DOI:10.3321/j.issn:1672-9072.1999.11.020 |
史作民. 1998. 暖温带宝天曼植物群落种间联结及种群生态位//刘世荣, 蒋有绪, 史作民. 中国暖温带森林生物多样性研究. 北京: 中国科学技术出版社, 99-117
|
王伯荪, 彭少麟. 1985. 南亚热带常绿阔叶树种间联结测定技术研究. Ⅰ.种间联结测试的探讨与修正. 植物生态学与地植物学丛刊, 9(4): 272-285. |
Cox G W. 1972. 蒋有绪, 译. 普通生态学实验手册. 北京: 科学出版社, 34-35
|
Greig-Smith. 1983. Quantitative plant ecology. 3rd version. Oxford: Blackwell Science Publications, 105-112.
|
Jonsson B G, Moen J. 1998. Patterns in species associations in plant communities: the importance of scale. Journal of Vegetation Science, 9: 327-332. DOI:10.2307/3237097 |