 2006, Vol. 42
2006, Vol. 42文章信息
- 李六林, 季兰.
- Li Liulin, Ji Lan.
- 杂种榛子不同方位叶片光合作用的日变化
- Diurnal Variation in Photosynthesis of Differently Directional Leaves in Hybrid Hazels (Corylus heterophylla×Corylus avellana)
- 林业科学, 2006, 42(12): 47-53.
- Scientia Silvae Sinicae, 2006, 42(12): 47-53.
-
文章历史
- 收稿日期:2005-11-28
-
作者相关文章


榛子含有丰富的脂肪、蛋白质、碳水化合物及多种维生素和矿物质,具有很高的经济价值而倍受人们关注。我国有丰富的野生榛子资源,其中包括平榛(Corylus heterophylla)、毛榛(C. mandshurica)、刺榛(C. titetica)等12种(庞发虎等,2002),以平榛分布较多而且普遍。但平榛有果个小、果壳厚、出仁率低、产量不高等弱点,而从国外引进的大果欧榛果大、品质优,但抗寒性差,从而严重地限制了我国榛子的发展。光合作用是植物生长发育的基础和生产力高低的决定性因素,同时又是一个对环境条件变化很敏感的生理过程(许大全, 1992;Day et al., 1998;Franks et al., 1999)。植物叶片光合作用日变化是植物生产过程中物质积累与生理代谢的基本单元,也是分析环境因素影响植物生长和代谢的重要手段。为此, 众多研究者对枣(王永蕙等,1990)、苹果(张绍铃等,1991)、核桃(张志华,1993)、阿月浑子(路丙社等,1999)和猕猴桃(谢建国等,1999)等果树光合速率的日变化及其环境因子进行了研究,不仅掌握了这些果树生长结果与环境因子之间的关系,而且也为优质、高效栽培提供理论依据。近年来,我国选育的平欧杂种榛子不仅在果个大小、坚果品质、丰产性等方面均有了很大的提高,而且其抗寒性超过了国外欧榛品种。目前,平欧杂种榛子在国内许多地区进行了引种试验,并已初步掌握了优良品种的某些生物学特性(杨丽涛等,2000;孙振良等,1999)。本研究拟采用在黄土高原地区长势良好且未发生冻害的平欧110号和平欧349号品系的杂种榛子为试验材料,对其光合速率、气孔导度、胞间CO2浓度等生理指标和与其相关的生态指标进行测定和分析,以期进一步掌握平欧杂种榛子的生理生态特性,为其扩大栽培范围和栽培管理提供理论依据。
1 材料与方法试验于2003—2004两年的6—7月在山西农业大学园艺站榛子园内进行。该区位于山西省晋中市,属暖温带大陆性季风气候。年均气温8.9 ℃,极端最高温38.5 ℃,最低温-26.5 ℃。年均降水量500~600 mm,无霜期180 d;土壤母质为黄土,土层深厚。
试材为2~3年生的平欧110号和平欧349号(均为平榛和欧榛的杂交品种)榛子。试材于2002年春季栽植,定植的苗木均为根蘖苗,株行距为3 m×3 m,在生长期内除进行灌水、施肥等常规管理外,还在树冠下进行生物覆盖和生长季节根外施肥,以促进试材的健壮生长。选择树冠上部东、南、西、北4个方位开张角度相近的枝条,对其上第6~7节叶片进行分析测定(这些叶片的开张角度与枝条的开张角度基本一致,即这些叶片与它所着生的枝条是平行的)。于6—7月选择3个晴天,在5:00—19:00,使用美国CID公司生产的CI-301便携式光合测定仪,采用开放气路,每隔1 h测定一次净光合速率(Pn),同时测定气孔导度(Gs)和细胞间隙CO2浓度(Ci)等相关生理指标和光量子通量密度(PFD)、叶片温度(TL)、环境相对湿度(RH)和环境CO2浓度(Ca )等生态指标,每个品种每个方向选3个叶片, 每个叶片重复测定2次,并将RH转化为水汽饱和差(VPD)进行分析。在11:00,叶片均匀地用多层纱布逐渐遮光或用黑布完全遮光进行处理,待处理10 min后,测定PFD和Pn,以分析叶片的光饱和点和补偿点。
2 结果与分析 2.1 杂种榛子不同方位叶片光合速率的日进程及CO2同化量的比较 2.1.1 杂种榛子不同方位叶片光合速率的日变化杂种榛子Pn日变化如图 1所示,平欧110号南面叶片(叶片的正面朝北)的光合速率的日变化为双峰曲线,最大值出现在7:00,为10.05 μmol CO2·m-2s-1,此后Pn逐渐下降,于15:00出现低谷,而后回升于17:00出现第2次高峰,以后Pn逐渐下降。西面叶片(叶片的正面朝东)Pn的日变化为单峰曲线,高峰出现在8:00,然后逐渐下降,13:00以后一直维持在较低的水平;北面叶片(叶片的正面朝南)Pn的日变化也呈现双峰曲线变化,两高峰分别出现在10:00和14:00—15:00;东面叶片(叶片的正面朝西)Pn从5:00开始逐渐升高,6:00—18:00 Pn一直维持在一定的水平上,变化的范围3.22~4.92 μmol CO2·m-2s-1。

|
图 1 杂种榛不同方位叶片Pn、Gs、Ci的日变化 Fig. 1 Diurnal variation in Pn, Gs, Ci of differently directional leaves in hybrid hazels A、C、E分别是平欧110号Pn、Gs、Ci的日变化;B、D、F分别是平欧349号Pn、Gs、Ci的日变化;s:南面叶片;w:西面叶片;n:北面叶片;e:东面叶片。下同。 A, C and E are Diurnal variation in Pn, Gs and Ci of Ping-ou 110, respectively; B, D and F are Diurnal variation in Pn, Gs and Ci of Ping-ou 349, respectively; s:Southern leaves; w:Western leaves; n:Northern leaves; e:Eastern leaves. The samebelow. |
南、北面叶片的Pn均为双峰曲线,且均为第1峰值较第2峰值高而且出现的时间长。但以上叶片Pn峰值出现的时间不同,上午第1峰值出现的先后顺序为南、北面的叶片,下午第2峰值出现的顺序正好相反。
平欧349号南面叶片和北面叶片的Pn变化规律与平欧110号基本一致(图 2),均呈双峰曲线,但第1高峰出现的时间有一定的差别;西面叶片Pn变化同平欧110号一样于7:00—8:00出现高峰,11:00后,Pn一直处于一个相对较低而稳定的水平;东面的叶片也同平欧110号基本一致,Pn的变化无明显的高峰和低谷。

|
图 2 不同方位叶片的PFD、TL、VPD和Ca的日变化 Fig. 2 Diurnal variation in PFD, TL, VPD and Ca of differently directional leaves |
平欧110号和平欧349号叶片的平均光合速率均以南面叶片为最高(表 1),依次为西面叶片、北面叶片,以东面叶片为最低。
|
|
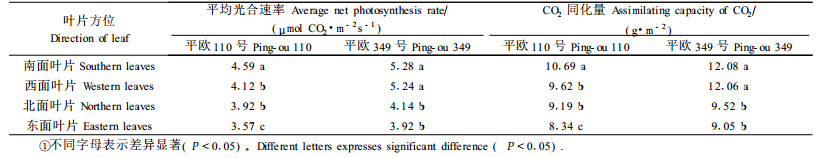
平欧110号南面叶片CO2日同化量为最高,分别是西面叶片、北面叶片的1.11和1.16倍,东面叶片CO2同化量为最低,仅相当与南面叶片的78%。平欧349号CO2同化量也以南面叶片为最高,是北面和东面叶片的1.27和1.33倍,与西面的叶片差异不明显。
2.2 杂种榛子光合速率与气孔导度和细胞间隙CO2浓度的关系 2.2.1 杂种榛子光合速率与气孔导度的关系平欧110号和平欧349号不同方位叶片气孔导度的日变化基本一致(图 1:C、D),从5:00开始,气孔导度逐渐增大,于8:00—10:00出现一天的最高峰,然后逐渐下降,有的虽然在17:00出现小高峰,但高值也较低。
相关分析结果表明:平欧110号和平欧349号南面叶片、西面叶片和北面叶片的Pn与Gs均呈极显著的正相关,而东面叶片的Pn与Gs无显著的相关关系。
2.2.2 杂种榛子光合速率与细胞间隙CO2浓度的关系平欧110号和平欧349号不同方位叶片细胞间隙CO2浓度的日变化也基本一致(图 1:E、F),从5:00开始,细胞间隙CO2浓度逐渐降低,于13:00—14:00出现增加并形成一个小高峰,然后逐渐下降,以后随着气孔的关闭和Pn的下降而迅速增加。
经相关分析表明,两品种的Pn与细胞间隙CO2浓度之间的相关系数均未达到显著水平,说明细胞间隙CO2供应不是导致Pn变化的直接原因,这与Farquhar等(1980)的观点相一致。
2.3 杂种榛子光合速率与环境因子分析 2.3.1 环境因子的日变化试验对叶片光合速率测定的同时,对影响光合作用的生态因子进行分析。结果发现影响两个品系光合速率环境因子的大小和变化基本相同,所以在论文中只列出了平欧110号不同方位叶片的变化。PFD日变化均为单峰曲线(图 2),但不同方位叶片高峰值出现的时间不同,南、西、北和东面叶片高峰出现的时间分别为12:00、10:00—11:00、12:00—13:00和13:00—15:00。不同方位叶片叶温的变化基本一致,早晨随光照强度的增强,温度不断上升,于13:00出现全天的最高温,以后一直保持在相当高的水平上,18:00以后温度才有所下降。
不同方位叶片周围空气水汽饱和差的大小和变化趋势基本一致。从早晨5:00开始,随PFD的逐渐增强,促进了叶片和土壤中水分的不断蒸发,从而使水汽饱和差减小,于6:00达到低谷,以后随PFD和温度的升高,水汽饱和差逐渐增大,于13:00—15:00达到高峰,而后又下降。
空气CO2浓度随植物不断进行光合作用而下降,中午13:00随着植物光合速率的下降,以及空气温度的升高,促进了呼吸作用,从而导致了空气中CO2浓度出现了一个小高峰;以后随着光合速率的的提高,空气中CO2浓度保持在一个较低的水平;6:00以后,随光合速率的下降,CO2浓度又开始上升。
2.3.2 杂种榛子光合速率与环境因子数量关系分析以PFD、TL、水汽饱和差和Ca为自变量,以Pn为因变量,用SAS系统进行多元逐步回归分析(陶俊等, 1999;李六林等, 2003)(表 2)。影响平欧110号和平欧349号南、西面叶片Pn的主导因子均为水汽饱和差和PFD;北面叶片除了受水汽饱和差和PFD外,还受到Ca的影响;东面叶片主要受到水汽饱和差和Ca因子的影响。根据以上分析,叶片Pn在中午前后以及在下午均较低可能主要由于水汽饱和差大幅度增加所致。
|
|

表 3拟合方程是通过函数模型y=A+Blnx (y指Pn,x指PFD)得到的,由此方程计算叶片的光补偿点PFD=e-A/B(μmol·m-2·s-1);由抛物线模型y=ax2+bx+c求得光饱和点(黄丛林等, 1996)。同一品种不同方位叶片的光饱和点有一定的差异,其中以西、北面叶片的光饱和点为较高,东、南叶片光饱和点则较低;叶片的光补偿点也表现出同样的规律。两品种的光饱和点差异较大,而光补偿点差异不十分明显。
|
|
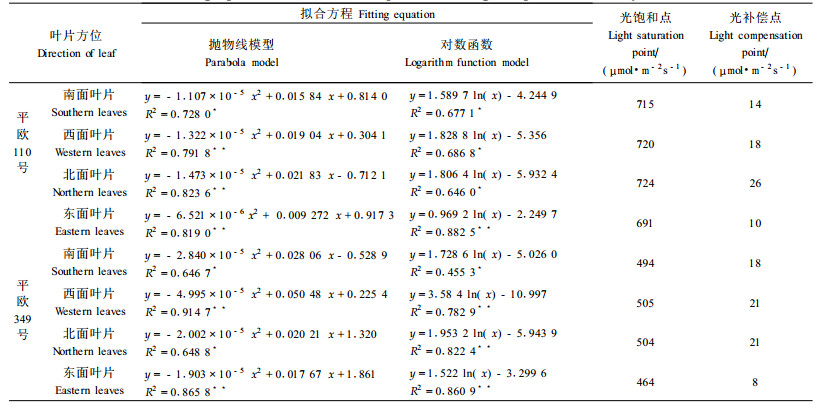
平欧110号和平欧349号的光饱和点分别约为691~724和460~505 μmol·m-2s-1,均较苹果(840 μmol·m-2s-1)(程来亮等,1992)、梨(1 100 μmol·m-2s-1)(王白坡等,1987)等和核桃(1 200 μmol·m-2s-1)(张志华等,1993)等果树低;其光补偿点分别约为10~26和8~21 μmol·m-2s-1,也同样低于苹果、梨和核桃等落叶果树。以上结果表明平欧110号和平欧349号对光照的要求较低,所以一般地区均可以满足杂种榛子对光照的要求,光照不为杂种榛子扩大栽植范围的限制因子。不同方位的光饱和点和光补偿点有一定的差异,这可能与其发育过程中适应不同的光照条件有关。平欧杂种榛是生长在我国东北地区的平榛和从欧洲引进的大果榛子的杂交种。平榛树体较小,在系统发育过程中长期处于森林群落内适应弱光或集生在荒山阴坡,所以对光照强度的要求,可能其光饱和点较低,而欧榛树体较大,一般生长在阳光充足的山坡,光饱和点较高(760~820μmol·m-2s-1,待发)。本试验供试的两个品系光饱和点差异较大,这可能是平欧110号在遗传特性上倾向于欧榛,所以其饱和点较高。而平欧349号则更倾向于平榛,所以其饱和点较低。
平欧110号和平欧349号不同方位叶片的微环境条件,除PFD外,TL、VPD和Ca的大小和变化规律基本一致。而其日平均光合速率和CO2同化量存在一定的差异,这可能主要由于PFD的不同所致。相比之下,南面叶片的PFD相对较低,但其日平均光合速率和CO2同化量却较高,这与叶片较低光饱和点的结论相一致,但在较高的PFD下,光合速率反而较低是否由于光抑制所致还需深入研究。
平欧110号和平欧349号在阳光充足、空气温度高、相对湿度小的夏季晴天,其南、北面叶片光合速率的日进程是一条双峰曲线,上下午各有个高峰,峰值的大小和出现时间有一定的差异,但均出现了明显的“午休”现象;西面叶片呈单峰曲线,峰值出现在上午,光合速率自中午降低后,一直处于较低的水平。许大全等(1990)、Xu等(1997)和沈允钢等(1998)提出,造成植物叶片净光合速午间降低的植物自身因素为气孔部分关闭引起的气孔限制和叶肉细胞自身活性下降引起的非气孔限制类,前者使胞间CO2浓度降低,而后者使胞间CO2浓度增高。研究结果表明:南、西、北叶片光合速率与气孔导度的相关性达到了显著或极显著的水平,午间这些叶片的气孔导度明显降低,但随着气孔导度的降低,细胞间隙CO2浓度呈现升高的趋势,说明午间光合速率降低不是主要由于气孔导度降低引起的,而是非气孔限制起着主要作用。至于西面叶片并没有像南北面叶片一样出现第二高峰,可能与下午光照强度较低有关。东面叶片的光合速率的日变化无明显的高峰,一直维持在一定的水平上,上午其光合速率没有出现高峰,主要与光照强度较弱有关;中午时,气孔导度明显降低,细胞间隙CO2浓度呈现升高的趋势,说明午间光合速率较低也主要受到非气孔的限制作用,因此尽管中午时光照强度有所增强,光合速率并没有增加。至于午后尽管光照强度较高,而光合速率并没有大幅度增加的原因还有待进一步研究。
对于大部分果树来说,其光合速率日变化规律是双峰曲线(王永蕙等,1990;张绍铃等,1991;路丙社等,1999;谢建国等,1999),在夏季,上午由于大气凉爽,随光照的增强,净光合强度也随之升高,10:00左右达最大值,之后,由于气温升高,光合强度随之逐渐降低,在14:00左右降至最低,甚至为负值,出现“午休”现象。在本试验中发现供试材料光合速率的高峰出现在7:00—8:00,较其他果树早,“午休”出现也较早。笔者认为平欧110号和平欧349号的光饱和点较其它低,在夏季早晨7:00—8:00的光强就接近饱和点,对影响净光合速率的环境因子进行逐步回归分析表明,VPD和PFD是影响光合速率的主导因子,而且VPD也较低,所以此时的条件适合榛子进行光合作用。Correia (1990)和郭连旺(1994)等认为光抑制和光呼吸的增强可能是引起植物叶片光合量子效率在中午降低的原因。强光、高温、低空气相对湿度、土壤干旱、气孔部分关闭等都可能是光合“午休”出现的原因。许多研究发现,空气湿度与“午休”也有密切联系,因为大多数植物的光合作用对空气湿度或确切地说对VPD很敏感(Raschke et al.,1986)。低空气湿度是引起“午睡”现象的重要生态因子(Beyschlag et al.,1987;娄长安等,1996),增加空气湿度可减轻“午睡”现象(娄长安等,1996)。试验中,9:00以后光照强度和气温快速增加,以及VPD增大,可能会诱发光抑制、光呼吸和暗呼吸升高,导致光合速率快速降低,以及前期光合效率高会不同程度地产生同化物积累的反馈抑制,所以导致了“午休”现象的提前。基于榛子光合的生理生态特点,在阳光充足的地区引种栽植时,可以适当加大栽培密度和幼树的行间可适当种植一些高秆的农作物,有条件的榛子园进行间歇喷灌,以减少水汽饱和差,改善生态环境条件,以提高幼树的光合产量和促进其健壮快速生长。
程来亮, 罗新书, 杨兴洪. 1992. 田间苹果光合速率日变化的研究. 园艺学报, 19(2): 111-116. |
郭连旺, 许大全, 沈允钢. 1994. 田间棉花叶片光合效率中午降低的原因. 植物生理学报, 20(3): 360-366. |
黄丛林, 张大鹏. 1996. 葡萄叶片光合速率日间降低内外因调控的研究. 园艺学报, 23(2): 128-132. |
李六林, 杨佩芳, 田彩芳, 等. 2003. 树莓光合特性的研究. 园艺学报, 30(3): 314-316. DOI:10.3321/j.issn:0513-353X.2003.03.015 |
娄长安, 莫庸, 王荣栋, 等. 1996. 新疆大陆性气候春小麦"午睡"现象的研究. 石河子农学院学报, 34(2): 1-5. |
路丙社, 白志英, 董源, 等. 1999. 阿月浑子光合特性及其影响因子的研究. 园艺学报, 26(5): 287-290. DOI:10.3321/j.issn:0513-353X.1999.05.002 |
庞发虎, 王勇, 杜俊杰. 2002. 榛子的特性及在我国的发展前景. 河北果树, (2): 1-2. DOI:10.3969/j.issn.1006-9402.2002.02.001 |
沈允钢, 施教耐, 许大全. 1989. 动态光合作用. 北京: 农业出版社, 55-60.
|
孙振良, 公建华, 张启昌, 等. 1999. 榛子、杂种欧榛、欧榛物候期及生长节律的初步调查. 吉林林学院学报, 15(1): 20-24. |
陶俊, 陈鹏, 佘旭东. 1999. 银杏光合特性的研究. 园艺学报, 26(3): 157-160. DOI:10.3321/j.issn:0513-353X.1999.03.004 |
王白坡, 丁兴翠, 戴文圣, 等. 1987. 田间条件下沙梨光合作用的研究. 园艺学报, 14(2): 97-102. |
王永蕙, 李保国. 1990. 枣树光合特性的研究. 华北农学报, 5(2): 65-70. DOI:10.3321/j.issn:1000-7091.1990.02.012 |
谢建国, 李嘉瑞, 赵江. 1999. 猕猴桃若干光合特性研究. 北方园艺, (2): 26-28. |
许大全. 1990. 光合作用"午睡"现象的生态、生理与生化. 植物生理学通讯, 26(6): 5-10. |
许大全. 1992. 植物光合作用的光抑制. 植物生理学通讯, 28(4): 237-243. |
杨丽涛, Timothy J. Arkebauer. 2000. 欧洲杨、榛子、短叶松和黑云杉气体交换的日变化. 植物生态学报, 24(4): 408-419. DOI:10.3321/j.issn:1005-264X.2000.04.005 |
张绍铃, 杨庆山, 马香莲, 等. 1991. 苹果短枝型品种光合特性研究. 果树科学, 8(3): 129-134. |
张志华, 高仪, 王文江. 1993. 核桃光合特性的研究. 园艺学报, 20(4): 319-323. |
Beyschlag W, Pfanz H. 1990. Stomatal Patchiness in sclerophyllous leaves. Plant Physiol, 93(suppl): 16. |
Correia M J, Chaves M Mc, Pereira J S. 1990. Afternoon depression in photosynthesis in grapevine leaves—evidence for high light stress effect. J Exp Bot, 41: 417-426. DOI:10.1093/jxb/41.4.417 |
Day W, Chalabi Z S. 1998. Use of models to investigate the link between the modification of photosynthesis characteristics and improved crop yields. Plant Physiol Biochem, 26: 511-517. |
Farquhar G D, Sharkeyr T D. 1980. Stomatal Conductance and Photosynthesis. Ann Rev Plant Physiol: 313-317. |
Franks P J, Farquhar G D. 1999. A relationship between humidity response, growth form and photosynthetic operating point in C3 plants. Plant Cell Envir: 1347-1349. |
Raschke K, Resimann A. 1986. The midday depression of CO2 assimilation in leaves of Arbutus unedo L: diurnal changes in photosynthetic capacity related to changes in temperature and humidity. Planta, 168: 546-558. DOI:10.1007/BF00392275 |
Xu Daquan, Shen Yungang. 1997. Diurnal variations in the photosynthetic efficiency in plants. Acta Phytophysiologica Sinica, 23(4): 410-416. |