 2006, Vol. 42
2006, Vol. 42文章信息
- 于晓英, 卢向阳, 姚觉, 邱收, 李达, 陈永华.
- Yu Xiaoying, Lu Xiangyang, Yao Jue, Qiu Shou, Li Da, Chen Yonghua.
- 瓜叶菊耐热无性系Z1-1-1的离体筛选与初步鉴定
- In Vitro Selection and Identification of Heat-Tolerant Clonal Line Z1-1-1 of Senecio cruentus
- 林业科学, 2006, 42(12): 33-38.
- Scientia Silvae Sinicae, 2006, 42(12): 33-38.
-
文章历史
- 收稿日期:2006-06-29
-
作者相关文章



2. 湖南农业大学生化与发酵工程实验室 长沙 410128;
3. 中南林业科技大学资源与环境学院 长沙 410004
2. Laboratory of Biochemistry and Fermentation Engineering, Hunan Agriculture University Changsha 410128;
3. College of Resources and Environment, Central South University of Forestry andTechnology Changsha 410004
瓜叶菊(Senecio cruentus)是菊科(Compositae)千里光属(Senecio)的一种观赏价值很高的多年生花卉(陈俊愉等,1989;包满珠,2003),花期正值少花的冬春季节(12月—翌年4月),花型秀美,花色艳丽,是冬春季节最常见的盆花,可供室内布置,温暖地区也可脱盆移栽露地布置早春花坛,还可用作花篮、花环的材料。但瓜叶菊不耐热,夏季育苗时往往遇到高温阻碍,幼苗死亡率很高(张建仪等, 1995;郭胜裕等,1996),所以生产中多作1、2年生栽培。改进其耐热性,对降低生产成本、实现周年供应优质产品及扩大其园林应用范围有重要意义。
体细胞无性系变异的离体筛选是现代生物技术中一个十分活跃的领域(Abdelbasset et al., 2005; Samantaray et al., 1999;2001; Mohamed et al., 2000; Svabovaet al., 2005; Peddaboina et al., 2005; Madhulika et al.,2003; Zairet al., 2003),通过高温直接筛选、抗羟脯氨酸(hydroxyproline,HYP)筛选、理化诱变高温胁迫筛选来获得耐热植物新品种,在乌菜(Brassica campestris)(陈静娴等,1995)、不结球大白菜(Brassica campestris ssp. chinensis)(黄剑华等,1995;王亦菲等,2002)、菜心(B. campestris spp. chinensis var. utilis) (何晓明等,1999;廖飞雄等,2004)、小麦(Triticum aestivum)(Mullarkwy et al., 2000)等多种作物上已有研究,对筛选出的变异进行抗性和生化指标的分析鉴定表明,这些变异体在热胁迫下相对电解质渗出率和丙二醛(MDA)含量都比较低,而同时有较高的脯氨酸含量,较好的细胞膜稳定性和较高的膜保护酶活性。离体培养在观赏植物快速繁殖上有很多报道,但在离体筛选特别是耐热品种筛选方面的研究报道却很少。虽然黄志刚等(2003)报道了非洲菊(Gerbera jamesonii)耐热变异离体筛选体系的研究,但到目前为止,还未见相关后续报道,同时也未见国内外有关瓜叶菊耐热无性系离体筛选的相关报道。为此,本试验在已建立的瓜叶菊离体培养和植株再生技术体系上,利用高温胁迫直接筛选、HYP与高温胁迫结合的方法进行了耐热无性系离体筛选的研究,并获得了1个耐热性比其母株有了很大提高的无性系,并对其再生苗移栽后相关特征进行初步鉴定。
1 材料和方法 1.1 植物材料用处于增殖期母株编号为B1、Z1的愈伤组织(切成约0.5 cm2的小块)和丛生芽(带3~5个小芽)为材料。材料的培养按于晓英等(2005)的方法进行。
1.2 方法 1.2.1 胁迫压的选择1) 热胁迫处理 将处于增殖期的愈伤组织(切成约0.5 cm2的小块)和丛生芽(带3~5个小芽)分别培养在恒温光照培养箱中,温度设为30、35、40 ℃,每天光照时数为10 h,光照强度为2 000 lx,设室温(17±2)℃作为对照。经不同时间高温处理后的材料再放于室温下进行恢复培养,21 d后统计存活率与增殖率,同时将能够恢复生长的进行继代培养。每个处理取10瓶,每瓶3~5个愈伤组织和丛生芽块,每个处理至少重复3次。
2) 渗透胁迫处理 选择羟脯氨酸(HYP)作为渗透胁迫选择剂,将愈伤组织和丛生芽分别培养在含有0、10、20、30、40、50 mmol·L-1 HYP的A4培养基中,配方见于晓英等(2005)。培养3周后,逐步将选择压去掉,统计去掉选择压培养21 d后的存活率、相对生长率和植株再生情况。每个处理取10瓶,每瓶4~5个愈伤组织和丛生芽块,每个处理至少重复3次。培养条件为每天光照时数为10 h,光照强度为2 000 lx,温度(17±2)℃。
3) 相对生长率的测定 相对生长量=(处理材料的终质量-始质量)/处理材料的始质量;相对生长率=(处理材料的相对生长量/未处理材料的相对生长量)×100%。
4) 存活率和增殖率的测定 材料没有完全变褐,仍有绿色或米黄色或乳白色的组织为存活个体。存活率=存活数/接种块数;增殖率=增殖芽数/接种丛芽数。
1.2.2 筛选方法直接选择:将继代培养1周左右处于增殖期的愈伤组织(米黄色或乳白色胚性愈伤组织)和丛生芽块,40 ℃胁迫下培养20 h,然后恢复到适温(17±2)℃,每天光照为10 h、光照强度为2 000 lx的环境下培养21 d。然后选取能够存活的愈伤组织和丛生芽块用同样的方法再筛选2~3代,用褐变作为伤害的判断指标。
HYP与温度胁迫结合:将继代培养1周左右处于增殖期的愈伤组织(米黄色或乳白色胚性愈伤组织)和丛生芽块,接种到含有30 mmol·L-1 HYP的A4培养基上;培养20 d之后,将能够恢复生长的绿色的愈伤组织和丛生芽块,接到含有20 mmol·L-1 HYP的A4培养基上适温(17±2)℃培养1周,再转入40 ℃胁迫下培养20 h;然后,再将能够恢复生长的愈伤组织和丛生芽块,在含有10 mmol·L-1 HYP的A4培养基上适温(17±2)℃恢复生长;21 d后,将能够增殖的愈伤组织和丛生芽块在40 ℃胁迫下再复筛1~2代。用褐变作为伤害的判断指标。
1.2.3 入选系植株再生将直接选择、温度与HYP胁迫结合筛选存活下来的愈伤组织和丛生芽,按照于晓英等(2004;2005)的方法进行增殖和根的诱导,使其形成完整植株(每瓶2~3株,共30瓶)。并将入选系的再生试管苗进行40 ℃(24 h)胁迫进一步进行耐热性的比较鉴定。
1.2.4 入选系移栽苗的初步鉴定将入选系及其对照母株生长健壮一致的再生苗移栽到装有相同基质的花盆中(各60株),在相同肥水管理和相同光照、温度条件下进行栽培养护。移栽30~40 d后检测相关指标。
气孔指数的测定参考刘艳菊等(2000)的方法:取叶片下表皮,在显微镜下观察单位面积(一个视野)范围内的气孔数。用以下公式表示:气孔指数=气孔数/(气孔数+表皮细胞数)。
热害指数的确定参考贾开志等(2005)的方法,将高温伤害程度划分为以下几级:0级,无热伤害症状;1级,少于1/4的叶片表现热伤害症状;2级, 1/4~1/2的叶片表现热伤害症状;3级, 1/2~3/4的叶片表现热伤害症状;4级, 3/4以上的叶片表现热伤害症状;5级,植株死亡。解除胁迫7 d后观察热害症状,按上述分级方法判定热害级数,通过以下公式计算热害指数:热害指数=∑各株级数/(最高级数×总株数)。
叶片细胞电解质伤害性外漏率(IL)和丙二醛(MDA)的测定参照王惠群等(1996)的方法,过氧化物酶(POD)和超氧化物歧化酶(SOD)活性的测定参考陈立松等(1997)的测定方法。
2 结果与分析 2.1 适宜选择压的确定 2.1.1 直接选择时适宜胁迫温度及胁迫时间的确定从表 1中可以看出B1、Z1的愈伤组织和丛生芽块的生长在35 ℃胁迫10 h后受到的抑制作用很小,存活率均≥96%,胁迫66 h时,存活率都开始急骤下降,到胁迫94 h时,来自母株编号为B1的材料存活率已降至0%;40 ℃恒温胁迫8 h时, B1、Z1愈伤组织的存活率都在90%以上,随着胁迫时间延长,处理材料的存活率开始快速下降,其存活率的差距也开始拉大,到24 h时,来自B1的存活率已为0.00%。解除胁迫后,愈伤组织恢复能力明显低于丛生芽块,并且热处理时间越长,恢复能力越差,相对生长率越低,新芽分化越迟,数目越少。
|
|
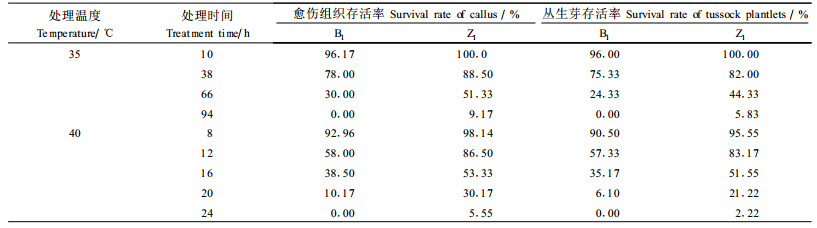
比较35 ℃和40 ℃热胁迫处理,后者选择压力较大,使材料到半致死的时间比较短,既节省了时间,也避免了因胁迫压低、培养时间长而出现的生理适应。所以为了筛选的效率与便利,为了能获得真正的耐热系,选出的耐热材料能更符合生产要求,认为40 ℃ 16~20 h胁迫处理可以作为瓜叶菊耐热变异无性系离体筛选的适宜温度和时间,处理后培养材料的存活率可以作为离体筛选的指标。
2.1.2 间接选择时适宜HYP浓度的确定试验中观察到在含有10 mmol·L-1和20 mmol·L-1HYP的培养基中培养3周后B1、Z1的愈伤组织仍有很高的存活率(>94%);40 mmol·L-1 HYP培养1周后,愈伤组织的平均存活率下降到了52.7%,培养3周后,大部分愈伤组织发生褐化,平均存活率急剧下降到16.1%;在50 mmol·L-1 HYP中培养1周后,大量的愈伤组织出现褐化,3周后平均愈伤组织存活率为0%。从图 1中也可以看出在含有10 mmol·L-1和20 mmol·L-1 HYP的培养基中培养20 d时,Z1愈伤组织的相对生长率比较高(分别为85.5%和78.5%);在含有30 mmol·L-1 HYP的培养基中培养12 d内Z1相对生长率增长缓慢,但在培养12 d后有快速增加;在含有40~50 mmol·L-1 HYP培养基中培养2周后,因大部分愈伤组织出现褐化,Z1的相对生长率没有增加,而是出现下降趋势。表明Z1的愈伤组织对低浓度的HYP(10~20 mmol·L-1)有较强的适应性,而40~50 mmol·L-1HYP渗透胁迫对Z1愈伤组织生长分化抑制作用很强。比较不同浓度HYP对瓜叶菊愈伤组织存活率及生长与分化的影响,同时也为了提高筛选的效率,维持一定数量的选择群体,认为30 mmol·L-1HYP可以作为瓜叶菊离体筛选的适宜浓度。

|
图 1 HYP对Z1愈伤组织相对生长率的影响 Fig. 1 Effect of HYP on the relative growth rate of callus of Z1 |
经过40 ℃ 20 h胁迫第1次筛选后,B1、Z1的大部分愈伤组织和丛生芽都变褐死亡,只有一小部分仍存活,其中愈伤组织存活率分别为11.4%和30.0%,丛生芽存活率分别为6.7%和22.1%。40 ℃ 20 h胁迫第2次筛选时,来自B1的愈伤组织和丛生芽全部受到伤害而变褐死亡,存活率为0%,而来自Z1的材料仍有少部分(11.1%)存活。40 ℃ 20 h胁迫第3次筛选时,Z1的丛生芽块和愈伤组织各只有1个存活,把这2个材料分别编号为Z1-1-1、Z1-1-2(图版Ⅰ-1, 2)。

|
图版Ⅰ Plate Ⅰ |
经过30 mmol·L-1HYP第1次筛选后,B1、Z1的愈伤组织存活率分别为32.6%和43.5%,丛生芽存活率分别为29.8%和38.9%。20 mmol·L-1HYP和40 ℃ 20 h第2次筛选时,B1、Z1的愈伤组织存活率分别降为10.9%和19.6%,丛生芽存活率分别为9.6%和15.6%。10 mmol·L-1HYP和40 ℃ 20 h第3次筛选时,来自B1母株的材料全部变褐死亡,存活率为0%;来自Z1的丛生芽和愈伤组织还有小部分存活,存活率分别为6.7%、9.8%。40 ℃ 20 h第4次筛选时,除Z1的愈伤组织还有1块存活外,其他材料已全部变褐死亡,把这个存活的材料编号为Z1-2-1 (图版Ⅰ-3)。
2.4 入选系植株再生将上述筛选中存活下来的Z1-1-1、Z1-1-2、Z1-2-1按照于晓英等(2004;2005)的方法进行增殖及芽的分化和根的诱导培养,但在培养过程中Z1-1-2没有获得再生植株。
2.5 入选系再生试管苗耐热性比较将经过增殖及芽和根的分化培养而获得的Z1-1-1、Z1-2-1离体再生苗,在40 ℃下持续热胁迫24 h,然后恢复到适温(17士2)℃培养20 d,进行耐热性的初步试验。结果发现在胁迫解除20 d后,入选系Z1-1-1的存活率高达63.3%(图版Ⅰ-6),而另一个入选系Z1-2-1和它的母株Z1已全部受伤致死(图版Ⅰ-4, 5)。
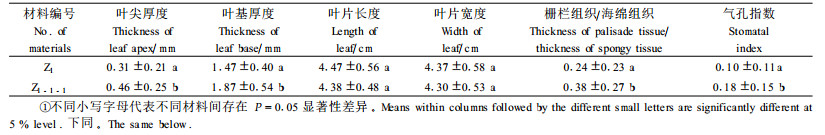
2.6 耐热无性系移栽苗的初步鉴定 2.6.1 在适温(14~22 ℃)下的性状表现将无性系Z1-1-1及其对照母株Z1的试管苗移栽到花盆中,相同条件下培养30 d后,开始观察其形态特征(表 2)。从表 2可知,无性系Z1-1-1和其母株Z1在叶片大小方面基本一致,但其叶片厚度、气孔指数、栅栏组织/海绵组织比值(TPT/TST)明显大于其母株Z1,其栅栏组织不仅比母株厚,而且排列也更整齐致密。
|
|
正常栽培管理条件下,进入花期后,无性系Z1-1-1和母株Z1在花色、花径大小、开花时间方面没有什么差别(图版Ⅰ-7, 10),但在花的育性上出现了较大的差异。Z1进入盛花期时,在筒状花上有很多金黄色的花粉(图版Ⅰ-8, 9),结实率85%以上。而Z1-1-1从始花到谢花期,一直未看到有花粉散出(图版Ⅰ-11),但剥开花药后可看到有花粉,只是花粉的畸形率高达40%(图版Ⅰ-12)。给Z1-1-1人工授予Z1的花粉后,Z1-1-1的结实率也仅1.0%。
2.6.2 耐热变异无性系在热胁迫下的性状表现在解除热胁迫(40 ℃ 24 h)7 d后统计各材料的热害程度如表 3,从表中可以看出经过高温处理后,无性系Z1-1-1的热害指数远远低于其对照母株,表现出了较强的耐热性,和其对照之间存在显著差异。来自Z1的植株只有极少数在解除胁迫、叶面喷水后1周能恢复正常生长,而来自变异Z1-1-1的植株几乎看不到很明显的伤害症状(图版Ⅰ-13~18)。
|
|
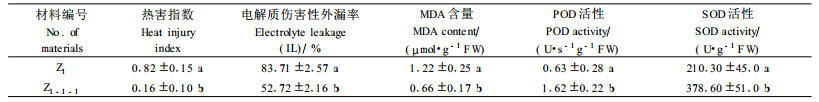
当40 ℃胁迫24 h结束后,立即测定Z1-1-1和其对照母株叶片中的电解质伤害性外漏率IL值、MDA含量、SOD和POD活性(表 3)。相对电解质渗出率IL和丙二醛含量反映出逆境下细胞膜的稳定性,从表 3可见,热胁迫后Z1-1-1的IL值明显低于其母株,说明这个无性系在热胁迫后,能使细胞膜保持较好的稳定性,防止电解质的渗漏。Z1-1-1的细胞丙二醛(MDA)含量同样比其母株低,差异显著,说明受胁迫后Z1-1-1的细胞膜过氧化程度远低于Z1,结果与电解质伤害性外漏率IL值一致。另外,细胞保护酶活性在一定程度上也反映出Z1-1-1抗性的增强。Z1-1-1的SOD活性(378.6 U·g-1FW)和POD活性(1.621 U·s-1g-1FW)也明显高于Z1的SOD和POD活性(分别为210.3 U·g-1FW和0.632 U·s-1g-1FW),存在显著差异。
3 讨论植物离体筛选技术是植物抗逆育种的一种快速有效的好方法,建立正确有效的分离筛选技术,是分离获得为数很少的所需突变细胞的保证。不同起始强度选择压处理会对选择后代的存活入选及稳定有一定影响,起始选择压低,前期淘汰率较低,需经较多的筛选世代和大量供选材料,不易漏掉本来就存在的一些变异,缺点是容易使生理适应性细胞存活下来;起始选择压高,前期淘汰率较大,筛选周期短,可能会在较短的时间内获得稳定的抗性系,缺点是可能把一些本来存在的可筛选出的抗性变异细胞淘汰,不利于捕捉到微小的变异类型。对于所采用的选择材料来源,有人认为体细胞诱导的愈伤组织和不定芽易出现嵌合体,实际上愈伤组织和不定芽再生的植株通常并不都是嵌合的,通过增殖培养,所出现的嵌合体常常会被逐步分离纯化。黄剑华等(1995)就用不结球大白菜幼苗为外植体诱导的增殖阶段的不定芽进行高温直接筛选获得了比原始材料更为耐热的变异体。廖飞雄等(2003;2004)以体细胞诱导的愈伤组织为材料,用羟脯氨酸为选择压,用高浓度一步筛选法,经3个周期筛选后获得了菜心耐羟脯氨酸初选系,经鉴定耐羟脯氨酸初选系具有较强的耐热性。但用愈伤组织进行高起始选择压处理时,往往由于高强度选择压所造成的胁迫伤害作用而丧失胚胎发生能力,较难获得再生植株。如赵宇玮等(2005)用一步正筛选法获得的5个耐甘露醇变异系中只有1个变异系保持了胚性状态。
本试验选取瓜叶菊体细胞诱导处于增殖阶段的愈伤组织和丛生芽为材料,通过对热胁迫、渗透胁迫处理后存活率、相对生长量、植株再生情况的分析比较,认为以丛生芽为材料植株再生比较容易,只是对上述几种逆境胁迫的忍受能力有限,存活率相对较低;而愈伤组织虽然在上述几种逆境胁迫下存活率高于丛生芽,但经过高温胁迫后再生植株比较困难。本试验中经过直接选择获得的入选系Z1-1-2就没有获得再生植株。但经过适当浓度(10~30 mmol·L-1)HYP处理后,愈伤组织的再生能力反而增强。试验中经过HYP与温度胁迫结合4次筛选获得的入选系Z1-2-1的愈伤组织成功分化出了再生苗,但再生试管苗耐热性并没有明显提高,在40 ℃温度下持续热胁迫24 h后受伤死亡,估计Z1-2-1可能只是一种逆境适应表现。
热胁迫下的存活率、生长量和膜透性等是植物耐热性鉴定和生理研究中最常用的指标,从这几个指标来看,本试验通过离体筛选技术初步选出的瓜叶菊耐热无性系Z1-1-1在热胁迫下有较低的相对电解质渗出率和MDA累积量,较高的SOD和POD保护酶活性,这些指标间接反映出其有较高的耐热性。在模拟高温下其再生移栽苗较低的热害指数直接反映出它有较强的耐热性且证实这种耐热性已经可以在无性繁殖过程中传递下去,但这种耐热性能否通过有性繁殖遗传给后代,则需要进一步加以研究。另外,试验只观察了Z1-1-130~60 d的移栽苗能耐7—8月夏季高温性状,至于移栽小于30 d或大于60 d的移栽苗是否可以忍耐7—8月夏季高温或更长时间的高温还有待进一步的研究。
另外,与母株Z1相比,无性系Z1-1-1除了耐热性得到了很大的提高外,在外部形态特征上也有较大的变异,表现出叶色浓绿,叶片变厚,花的育性降低,其结实率几乎为0。这些变异是由于染色体数目变化所致还是染色体结构变异或者是其他多基因的突变结果有待进一步的研究和探讨。
包满珠. 2003. 花卉学. 北京: 中国农业出版社, 181-182.
|
陈俊愉, 程绪珂. 1989. 中国花经. 上海: 上海文化出版社, 549.
|
陈静娴, 聂凡. 1995. 乌菜组织培养及耐热变异体的诱导和筛选. 安徽农业科学, 23(3): 201-205. |
陈立松, 刘星辉. 1997. 植物抗热性鉴定指标的种类. 干旱地区农业研究, 15(4): 72-77. DOI:10.3321/j.issn:1000-7601.1997.04.015 |
郭胜裕, 朱兵, 曹爱东, 等. 1996. 瓜叶菊高温期育苗技术. 河南农业科学, (8): 27. |
何晓明, 潘瑞炽, 廖飞雄. 1999. 菜心耐热变异体的离体筛选研究. 广东农业科学, (5): 17-18. |
黄志刚, 陈兆平, 文方德. 2003. 非洲菊耐热变异离体筛选体系的研究. 亚热带植物科学, 32(4): 25-29. DOI:10.3969/j.issn.1009-7791.2003.04.008 |
黄剑华, 陆瑞菊, 张玉华. 1995. 应用离体培养技术鉴定不结球大白菜耐热性及诱导耐热变异体. 上海农业学报, 11(4): 18-22. |
贾开志, 陈贵林. 2005. 高温胁迫下不同茄子品种幼苗耐热性研究. 生态学杂志, 24(1): 398-401. |
廖飞雄, 潘瑞炽. 2003. 菜心耐羟脯氨酸变异筛选方法的研究. 江西农业大学学报, 25(6): 875-878. DOI:10.3969/j.issn.1000-2286.2003.06.017 |
廖飞雄, 潘瑞炽. 2004. 菜心耐羟脯氨酸初选系的耐热性. 热带亚热带植物学报, 12(4): 359-362. DOI:10.3969/j.issn.1005-3395.2004.04.013 |
刘艳菊, 李承森. 2000. 中国辽宁省始新世水杉的研究. 植物学报, 42(8): 873-878. DOI:10.3321/j.issn:1672-9072.2000.08.016 |
王惠群, 孙福增, 彭克勤, 等. 1996. 淹水处理对水稻叶片膜脂过氧化作用及细胞膜透性的影响. 湖南农业大学学报, 22(3): 222-224. |
王亦菲, 陆瑞菊, 周润梅, 等. 2002. 以不结球白菜无菌苗茎尖诱导筛选耐热变异体. 上海农业学报, 18(2): 6-9. DOI:10.3969/j.issn.1000-3924.2002.02.002 |
于晓英, 卢向阳, 龙岳林, 等. 2005. 瓜叶菊花梗和花托愈伤组织的诱导与植株再生研究. 湖南农业大学学报, 31(4): 399-401. |
于晓英, 卢向阳, 吴铁明, 等. 2004. 瓜叶菊的离体培养与快速繁殖. 湖南农业大学学报, 30(5): 429-433. |
张建仪, 杜猛军, 许晓华, 等. 1995. 瓜叶菊高山育苗对调节花期的影响. 浙江农业科学, (6): 302-304. |
赵宇玮, 郝建国, 步怀宇, 等. 2005. 小麦耐甘露醇变异细胞系的离体筛选及植株再生. 西北大学学报:自然科学版, 35(2): 184-186. |
Abdelbasset El Hadrami, Abdelmalek El. 2005. Toxin-based in-vitro selection and its potential application to date palm for resistance to the bayoud Fusarium wilt. Comptes Rendus Biologies, 328(8): 732-744. DOI:10.1016/j.crvi.2005.05.007 |
Madhulika Singh, Uma Jaiswal, Vijai Shanker Jaiswal. 2003. In vitro selection of NaCl-tolerant callus lines and regeneration of plantlets in a bamboo (Dendrocalamus strictus Nees). In Vitro Cellular & Development Biology, 39(1): 229-233. |
Mohamed M A H, Henderson P J C. 2000. In vitro selection and characterization of a drought tolerant clone of Tagetes minuta. Plant Science, 159(2): 213-222. DOI:10.1016/S0168-9452(00)00339-3 |
Mullarkwy M, Jones P. 2000. Isolation and analysis of thermotolerant mutants of wheat. Journal of Experimental Botany, 342(5): 139-146. |
Peddaboina Venkataiah, Thamidala Christopher, Subhash Karampuri. 2005. Selection of atrazine-resistant plants by in vitro mutagenesis in pepper(Capsicum annuum). Plant Cell, Tissue and Organ Culture, 83(1): 75-82. DOI:10.1007/s11240-005-2678-4 |
Samantaray S, Rout G R. 2001. Induction, selection and characterization of Cr and Ni-tolerant cell lines of Echinochloa colona (L.) Link in vitro. Journal of Plant Physiology, 158(1): 1281-1290. |
Samantaray S, Rout G R, Das P. 1999. In vitro selection and regeneration of zinc tolerant calli from Setaria italica L. Plant Science, 143(2): 201-209. DOI:10.1016/S0168-9452(99)00036-9 |
Svabova L, Lebeda A. 2005. In vitro selection for improved plant resistance to toxin-producing pathogens. Journal of Phytopathology, 153(1): 52-64. DOI:10.1111/j.1439-0434.2004.00928.x |
Zair I, Chlyah A, Sabounji K, et al. 2003. Salt tolerance improvement in some wheat cultivars after application of in vitro selection pressure. Plant Cell, Tissue and Organ Culture, 73(1): 237-244. |