 2006, Vol. 42
2006, Vol. 42文章信息
- 刘雪梅, 杨传平.
- Liu Xuemei, Yang Chuanping.
- 白桦雌雄花发育周期的时序特征
- Temporal Characteristics of Developmental Cycles of Female and Male Flowers in Betula platyphylla in Northeastern China
- 林业科学, 2006, 42(12): 28-32.
- Scientia Silvae Sinicae, 2006, 42(12): 28-32.
-
文章历史
- 收稿日期:2006-06-12
-
作者相关文章



白桦(Betula platyphylla)是我国东北地区主要的阔叶树种。陈之端等(陈之端,1991;1994a;1994b;陈之端等,1995;2001)已对桦木科(Betulaceae)各属植物进行了全面而系统的系统发育研究。目前桦木科植物有关花及胚胎发育的报道中,主要有美洲铁木(Ostrya virginiana)的胚珠和胚囊发育(邢树平等,1998),鹅耳枥(Carpinus turczaninowii)和虎榛子(Ostryopsis davidiana)雌性生殖器官的形态发生(陈之端等,2001),白桦雌雄花胚胎发育的解剖学观察(刘雪梅等,1998;2005a;2005b;姜静等,2001)等,这些研究主要着重于胚胎学观察和系统发育研究。在白桦花发育周期、分子机理及基因调控方面,只有极少相关报道(杨传平等,2006),其主要原因是白桦的有性生殖周期较长,而花发育的关键阶段历时短,不易把握取材时机,而且白桦的花很小,给相应的实验操作带来很大困难。目前,对白桦花发育周期和时序特点尚缺乏报道。本文在白桦胚胎学研究的基础上(刘雪梅等,1998;2005a;2005b),研究白桦雌雄花在生殖发育周期和发育时序上的特殊性,以充实桦木属(Betula)植物花发育研究资料,并为白桦的有性繁殖提供科学依据。
1 材料与方法白桦的雌雄花序于2001—2002年采自东北林业大学校园内20~30年生白桦植株,于FAA固定液中固定和保存。用OLYMPUS显微镜进行实体解剖观察,同时采用石蜡生物制片技术制片,爱氏苏木精或铁矾苏木精染色,利用ZEISS-A xioskop2型显微镜观察和摄影。
2 结果与分析 2.1 白桦雌雄花的结构特征观察结果表明,白桦花单性,雌雄同株。白桦雄花序多分布在树冠中下部,2或3个并生或单生于当年生枝条的顶端。雌花序为圆柱形,多着生在树冠中下部的1年生以上的长枝或短枝上。实体解剖和显微制片观察结果表明,雌花序外被许多大苞片,每苞腋内有3朵雌花和2小苞片,雌花无花被,具合生心皮雌蕊。子房具两室,绝大多数每室1个胚珠(图版Ⅰ-1,2),但也发现极个别为每室有2个胚珠(图版Ⅰ-3)或没有胚珠。中轴胎座,每个雌蕊具有2个花柱;胚珠倒生(图版Ⅰ-1,2),单层珠被,厚珠心发育(图版Ⅰ-4),蓼型胚囊。雄花序为下垂的柔荑花序,常先叶开放。每苞腋内有3朵小花,花丝极短,每花具2个雄蕊,花药基着生,每个花药具2个药室,2个花粉囊基部结合于花丝(图版Ⅰ-5,6)。

|
图版Ⅰ 1.胚珠发育初期,显示白桦子房具二室,每室具1个胚珠,胚珠倒生(×100);2.胚珠发育中期,显示子房具二室,每室具1个胚珠,胚珠倒生(×90);3.有的子房室具2个胚珠(×100);4.二核胚囊,显示厚珠心胚珠(×500);5.花药发育初期,显示每个雄花含2个雄蕊,显示每个花药具2个药室初始细胞团(×120);6.小孢子四分体时期,显示每个花药具2个药室(×100);7.左边为二核胚囊,右边为二核分裂中期(×100);8.2个胚珠发育不一致,其中右边的胚珠有退化趋势(×80);9.二细胞原胚时,另1个胚珠已退化(×100) Plate Ⅰ 1. At the initial stage of ovule development, wary has 2 locules, with 1 ovule in each locule and anatropous ovule (×100); 2. At the middle stage of ovule development, ovary has 2 locules, with 1 ovule in each locule and anatropous ovule (×90); 3. Two ovules in some locule (×100); 4. Two-nucleus embryo sac and cmssinucellate ovule (×500); 5. At the initial stage of anther development, each male flower contains 2 stamens, and each anther has 2 anther cells f original cell groups (×120); 6. At the tetrad stage of micrspore, each anther has 2 anther cells (×100); 7. Two-nucleus e mbryo sac (left), metaphase of two-nucleus division (right) (×100); 8. Two ovules of developments are different, and the right ovule has a trend of degeneration (×80); 9. At the two-cell prcembryo stage, the oher ovule has degenerated (×100) |
雌花芽于6月初出现花序原基;6月中旬,花序原基上产生大苞片原基;7月中旬,大苞片原基基部出现子房原基;9月中旬,在大苞片和子房原基之间从基部产生小苞片原基,并以此状态越冬,直到第2年4月20日左右从花芽中露出雌花序。
4月24—26日雄花开始大量散粉,并持续1周左右,这期间雌花的柱头成熟,开始接受花粉,但此时胚囊还未发育成熟,因此不能立即受精,受精是在受粉之后约1个月左右进行。这1个月中,一方面,花粉管沿柱头向胚囊方向生长,或者二核花粉粒还可能再进行一次有丝分裂;另一方面,子房和胚珠继续发育,胚囊内逐次进行大孢子发生和雌配子体形成。5月15—18日,单层珠被的原基已形成,珠心组织内出现孢原细胞、大孢子母细胞,还有的材料此时减数第1次分裂已经结束,为二孢子时期,5月20日左右,少数已发育为直线型大孢子四分体;5月21—24日期间,合点端的功能大孢子进行1次有丝分裂,形成二核胚囊,珠孔端与合点端各有1个核,二核胚囊继续发育为四核胚囊,两极各有2个核;5月25 —28日则既有在胚囊两极各平均分配四核的八核胚囊时期,又有成熟胚囊时期。5月29日左右,胚囊已完全成熟,此时可进行受精。白桦大孢子发生和雌配子体发育过程表明,白桦属于典型的单孢子蓼型胚囊发育类型。据文献报道,在研究过的有花植物中,有70%属于此类(胡适宜,1982)。
5月28—30日进行受精作用,当产生雄性小核仁时,周围已产生32个游离核。可见,精子与2个极核或次生核的融合早于精子与卵细胞的融合。由于融合后的初生胚乳核经过有丝分裂产生许多游离核,因此,白桦的胚乳类型为核型胚乳。5月30日—6月1日,形成初生胚乳核(或次生核)和受精卵。此后,进入胚和胚乳发育时期。6月2—8日为原胚发育阶段,前后依次经历二细胞原胚、三细胞原胚、球胚、心形胚等阶段,而胚乳则由游离胚乳核发育为细胞型胚乳;6月9—12日形成鱼雷胚,6月15—19日鱼雷胚体积明显增大,周围的胚乳已开始被吸收,7月5日以后继续胚的发育,直到8月中下旬,胚发育成熟,此时,种子成熟。
2.3 白桦雄花发育的时序特点5月30日左右雄花序露出芽鳞,开始雄花发育。7月中旬至8月中旬是小孢子发生和雄配子体形成时期。7月10—15日,出现孢原细胞,7月15—20日,发育为小孢子母细胞;7月19—28日的材料中可见到处于细胞分裂的前期Ⅰ、中期Ⅰ、后期Ⅰ的细胞;7月28日—8月5日期间多数细胞处于后期Ⅰ和末期Ⅰ状态。至此,减数分裂的第1次分裂到此结束,但2个子核之间不形成细胞壁,且外被胼胝质壁。
减数分裂的第2次分裂发生于8月6—13日,可看到前期Ⅱ、中期Ⅱ、后期Ⅱ和末期Ⅱ;8月13—14日,减数分裂形成的4个小孢子仍包被在同一胼胝质壁中,形成形态比较固定的四分体类型。8月14—16日,四分体解体,小孢子在药室中处于游离状态,每个小孢子均处于单核时期,并以此状态越冬,这时,小孢子可明显看到3个萌发孔。第2年的4月中旬,单核小孢子开始继续发育,进行有丝分裂,4月下旬—5月初形成具二核或三核的成熟花粉,并进行散粉。二核花粉以后在花粉管中再进行1次有丝分裂形成三核花粉。
上述白桦雌雄花发育周期列于表 1。需要指出的是,此表中的时间和发育状态不是绝对的,即在同一花序内,其中各个小花的发育状态也不是一致的,例如白桦同一子房内不同的2个胚珠中1个胚珠内为二核胚囊期,另1个胚珠中的二核正处于向四核胚囊分裂的中期(图版Ⅰ-7);或1个胚珠在成熟胚囊受精后继续胚胎发育,但另1个胚珠却逐渐退化(图版Ⅰ-8,9)。种实成熟时,只是其中的1个胚珠发育为成熟胚,另1个完全退化萎缩。
|
|
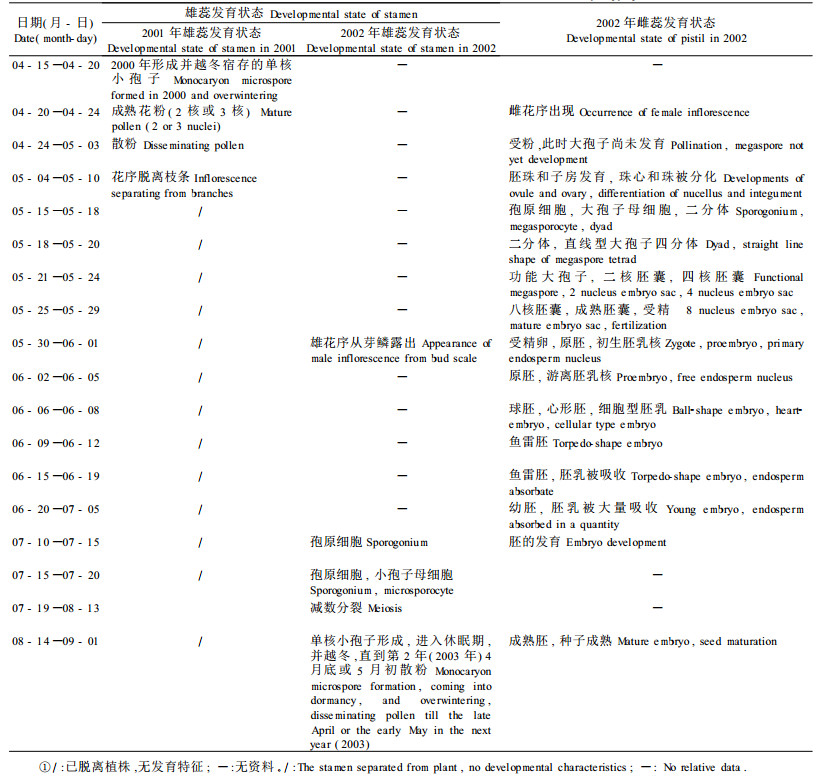
植物的有性生殖是雌花与雄花高度有序、相互平衡发育的结果。雌花和雄花不仅在结构和功能上表现出适应性,二者的发育进程也表现为协调一致。不同种的植物花发育时间是不同的,除了自身的遗传因素外,很大程度上也取决于植物的生长环境。不仅如此,同种植物的雌花和雄花发育在时间上也存在不一致性。白桦的有性生殖周期较特殊,表现出雌花和雄花发育程度的差异(表 1)。
由表 1可见,白桦的雌雄花发育周期有以下特点:1)花序出现及减数分裂时间不同。雌花序出现于4月中下旬,雄花序出现于5月底,二者相差近1个月。大孢子母细胞于5月中旬开始减数分裂,历时3~4 d,而小孢子母细胞于7月中下旬开始减数分裂,8月中旬结束,历时1个月。2)雌雄配子体的成熟历时不同,白桦的雄配子体发育很特殊,雄花的花粉发育历经2年,其间包括休眠期。一般来说,小孢子出现于8月13日,但是,当年不能发育为成熟的雄配子体,而仅仅以单核小孢子状态宿存于雄花序中越冬,直到第2年4月底至5月初才发育成熟,形成具有二核或三核的成熟花粉,随即散粉。就是说,从小孢子至成熟雄配子体(花粉)需要历时8个月,期间休眠4~5个月。而雌花的大孢子和雌配子体发育当年完成。4)每年春天雌花所接受的花粉来源于前一年产生并在冬季宿存的雄花。虽然在4月底至5月初散粉时,当年生白桦雌雄花序在结构上能够保证有效的受粉,但此时胚囊还未成熟,不具备受精能力,直到20~30 d后胚囊发育成熟才可进行受精作用。
3 结论与讨论白桦雌花序出现于4月中下旬,大孢子母细胞于5月中旬开始减数分裂,历时3~4 d,5月20日左右形成大孢子,成熟胚囊在5月28日左右产生,之后进行受精和胚胎发育。雄花序出现于5月底,小孢子母细胞于7月中旬开始减数分裂,8月中旬结束,形成小孢子,但当年不再继续发育为成熟的雄配子体,而仅仅以单核小孢子状态宿存于雄花序中越冬,直到第2年4月底至5月初发育成熟,形成具有成熟花粉,然后散粉。所以,雌花所接收的花粉来源于前1年出现、宿存越冬、次年4月底开始散粉的雄花。
本研究结果表明,白桦花粉到达柱头后,雌配子体尚未发育成熟,需经历1个月后才完成受精。据文献记载,从花粉落到柱头上至雌雄核融合所经历的时间因植物的不同而异,有的可长达1年多,有的只有10多分钟(胡适宜,1982)。可见,不同植物的传粉和受精的间隔期,在一定的环境中是比较稳定的,主要与花粉管生长和胚珠结构有关。在植物体上生长的花粉管, 因要经多次重新取向, 才能进入胚囊, 具有明显的向性生长特性(邢树平,1998)。比较流行的是向化性假说和机械假说。前者认为由胚珠产生一种引导花粉管生长的信号,后者认为花粉管的向性生长主要依靠雌性生殖组织的结构和化学性质(Hulskamp et al., 1995)。菠菜(Spinacia oleracea)的花粉管在受精完成后呈吸器状,据说是受精胚珠释放的一种因子所致(Derksen et al., 1995)。Hulskamp等(1995)在拟南芥(Arabidopsis thaliana)突变体中的试验结果说明,胚珠作为一个自主功能单位,与花粉管的最后的向性生长有关,并进一步说明胚珠的孢子体组织对花粉管的向性生长可能不起作用,而真正起作用的是胚珠内的正常发育的雌配子体,即胚囊。可见,花粉与柱头的接触激发了某种物质产生或引起某些生化反应,促进了雌配子的发育。
作者在取材过程中观察到:白桦雌雄花在植株上的着生方式也很特殊,估计也与雌雄花花期不遇现象有关。白桦为雌雄同株异花植物,每年的4月底至5月初是哈尔滨市植物迅速复苏的时期,白桦的雌花序也在此时从雌花芽中露出,但雌花枝为一极短的短枝,在花序下方只有2个叶片,此短枝不存在枝条顶芽生长点,不存在枝条伸长的营养生长。而雄花序则相反,着生于具有旺盛营养生长枝条的顶端,所以雄花序要等到枝条伸长到一定程度,才会在枝条顶端的当年最后一个芽内萌发出来,此时已是5月底6月初,比雌花序的出现晚了约1个月,因此,呈现当年雌雄花的花期不遇现象。另外,雌花序发育较迅速,在雌花序出现后的1个月内(5月底)便完成了雌配子体的发育,并进入可受粉状态;而雄花序发育迟缓,直到雄花序出现后的1个月(7月初)才开始减数分裂和小孢子的发育。这种同年生的雌雄花严重异熟现象直接导致了二者不能受粉,雌花只能接受前一年生雄花的花粉。
本文结果已证实白桦的花粉发育历经2年,其间进行休眠越冬。这种现象在被子植物的阔叶树中是很罕见的(王宝娟等,2005)。通常在裸子植物中,小孢子及雄配子体发育常出现休眠现象,曹玉芳等(1995;1997)将其归纳为3种类型:1)小孢子母细胞经冬季休眠后,在春季解除休眠进入减数分裂期,如欧洲赤松(Pinus sylvestris)、欧洲云杉(Picea abies)和粉绿云杉(Picea glauca);2)在休眠以前,小孢子母细胞进行减数分裂形成成熟花粉粒,如落叶松(Larix gmelinii)、扁柏属(Chamaecyparis)和红豆杉属(Taxus)植物;3)减数分裂起始于休眠之前,休眠期间停止,解除休眠以后才完成发育,如欧洲落叶松(Larix decidua)、太白红杉(Larix chinensis)和花旗松(Pseudotsuga menziesii)的小孢子母细胞是以双线期渡过休眠,而异叶铁杉(Tsuga heterophylla)和北美乔柏(Thuja plicata)是以粗线期渡过休眠。以第3种类型中的太白红杉为例(俞晓敏等,2003),其小孢子母细胞在8月中旬形成以后进入减数分裂Ⅰ, 于10月上旬进入双线期;而10月中旬至次年的3月下旬近5个半月的时间是小孢子母细胞的休眠期,此休眠期解除后便进入减数分裂的前期Ⅰ的终变期,并继续发育至成熟花粉。Ekberg等(1968)认为, 假若落叶松属的某些植物在休眠期以前继续从双线期进一步发育, 将会降低其对严寒的抵抗力, 进而影响到正常花粉粒的形成。因此, 太白红杉以双线期渡过休眠, 应是其适应外界环境变化, 抵制严寒的一种机制(俞晓敏等,2003)。小孢子及雄配子体发育暂时中断现象,在其他阔叶树中并不多见,很少有报道(王宝娟等,20 05),例如,与白桦具有同样生活环境的水曲柳(Fraxinus mandshurica)的雌雄花是当年产生,雌雄配子体当年成熟,并受精结实,即当年生雌雄花在受粉和受精上保持较高一致性(詹亚光等,2005)。但白桦却存在此现象,当其减数分裂完成并形成单核小孢子时,已是8月中下旬,天气转冷,不再继续发育,而是在具有坚实苞片雄花序内休眠大约7个月(从8月中旬至次年的3月中旬),其中包括气温低达-30 ℃左右的严冬。从适应性和发育角度分析,白桦以单核小孢子方式越冬一方面能免受严寒侵袭,另一方面能保证次年春天其花粉的有效利用。类似白桦花粉的特殊发育方式的相关机理未见有系统报道,还有待于进一步研究。
曹玉芳, 许方, 姚敦义. 1995. 松杉类植物雌雄配子体的发育与胚胎发生. 莱阳农学院学报, 12(3): 206-212. |
曹玉芳, 姚敦义, 许方, 等. 1997. 侧柏小孢子的发生和雄配子体的形成. 西北植物学报, 17(2): 163-169. DOI:10.3321/j.issn:1000-4025.1997.02.006 |
陈之端. 1991. 桦木科植物的花粉形态研究. 植物分类学报, 29(6): 494-503. |
陈之端. 1994a. 桦木科植物的系统发育和地理分布. 植物分类学报, 32(1): 1-31. |
陈之端. 1994b. 桦木科植物的系统发育和地理分布:续. 植物分类学报, 32(1): 101-153. |
陈之端, 路安民. 1995. 桦木科植物的起源和早期分化. 中国科学院研究生院学报, 12(2): 199-204. |
陈之端, 邢树平, 梁汉兴, 等. 2001. 鹅耳枥和虎榛子(桦木科)雌性生殖器官的形态发生. 植物学报, 43(11): 1110-1114. DOI:10.3321/j.issn:1672-9072.2001.11.002 |
胡适宜. 1982. 被子植物胚胎学. source: 高等教育出版社, 1-182.
|
姜静, 杨传平, 刘桂丰, 等. 2001. 白桦雌雄花发育的解剖学观察. 东北林业大学学报, 29(6): 11-14. DOI:10.3969/j.issn.1000-5382.2001.06.004 |
刘雪梅, 杨传平. 2005a. 东北地区白桦雌配子体的形成与胚胎发育研究. 植物学通报, 22(2): 147-152. |
刘雪梅, 杨传平, 耿峰. 2005b. 白桦胚珠发育及胚、胚乳发育关系的研究. 植物研究, 25(3): 322-326. |
刘雪梅, 刘桂丰, 刘关君, 等. 1998. 白桦雄配子体的发育. 东北林业大学学报, 26(3): 38-41. |
王宝娟, 吉成均, 胡东. 2005. 银杏雄配子体发育的细胞学研究. 西北植物学报, 25(7): 1350-1356. DOI:10.3321/j.issn:1000-4025.2005.07.012 |
邢树平. 1998. 花粉管生长调控的研究进展. 广西植物, 18(1): 82-88. |
邢树平, 陈之端, 路安民. 1998. Ostrya virginiana (Betulaceae)的胚珠和胚囊发育及其系统学意义. 植物分类学报, 36(5): 428-435. |
杨传平, 魏继承, 李同华, 等. 2006. 实时定量PCR法分析白桦中一花发育相关基因BpHEN的时序表达. 东北林业大学学报, 34(1): 2-6. |
俞晓敏, 赵桂仿. 2003. 太白红杉小孢子的发生和雄配子体的发育. 植物学通报, 20(5): 576-584. DOI:10.3969/j.issn.1674-3466.2003.05.009 |
詹亚光, 尹立辉, 刘雪梅, 等. 2005. 水曲柳大小孢子发生、雌雄配子体发育及其系统学意义. 北京林业大学学报, 27(5): 42-47. DOI:10.3321/j.issn:1000-1522.2005.05.007 |
Ekberg G, Eriksson G, Sulikova Z. 1968. Meiosis and pollen formation in Larix. Hereditas, 59: 427-438. |
Hulskamp M, Schneitz K, Pruitt R E. 1995. Genetic evidence for a long-range activity that directs pollen tube guidance in Arabidopsis. Plant Cell, 7: 57-64. DOI:10.2307/3869837 |
Derksen J, Rutten T, Amstel T V, et al. 1995. Regulation of pollen tube growth. Acta Bot Neerl, 44(2): 93-119. DOI:10.1111/j.1438-8677.1995.tb00773.x |