 2006, Vol. 42
2006, Vol. 42文章信息
- 艾畅, 徐立安, 赖焕林, 黄敏仁, 王章荣.
- Ai Chang, Xu Li'an, Lai Huanlin, Huang Minren, Wang Zhangrong.
- 马尾松种子园的遗传多样性与父本分析
- Genetic Diversity and Paternity Analysis of a Seed Orchard in Pinus massoniana
- 林业科学, 2006, 42(11): 146-150.
- Scientia Silvae Sinicae, 2006, 42(11): 146-150.
-
文章历史
- 收稿日期:2005-10-25
-
作者相关文章



2. 国家林业局森林资源监督管理办公室 北京 100714
2. Office of Forestry Resources Supervision and Administration, the State Forestry Administration Beijing 100714
林木种批品质的优劣应该从3个方面来理解:第一, 播种品质, 包括种子饱满度、千粒质量、发芽率等指标; 第二, 遗传品质, 要求长成的个体具有优良的遗传特性, 如生长量、木材品质等; 第三, 由于种批的遗传多样性大小直接关系到造林群体的适应性、抗逆性与稳定性, 因此种批的遗传基础宽窄也应该作为种批品质的评价指标。当前, 在人工造林中, 种子通常来自于天然林或者是种子园, 绝大多数是自由授粉的, 而这些自由授粉的种子通常可能由固定的几个父本和母本交配产生(Lexer et al., 2000), 其遗传多样性将直接影响到人工林的稳定性和适应性。
亲本分析主要是指通过观察和(或)采用遗传标记研究个体或种群的亲缘关系(陈小勇, 1999), 从而为子代找到亲本。亲本分析包括父本分析和母本分析, 父本分析可以提供子代父本组成、花粉来源、自交等信息。本研究通过对种子园及其自由授粉子代的遗传多样性分析和父本分析, 探讨种子园自由授粉子代的遗传基础, 分析遗传多样性同父本组成之间的联系, 为利用种子园集约生产种子提供理论依据。
1 材料与方法 1.1 材料来源试验材料取自福建省五一林场马尾松种子园的东区东坡。东区共配置129个无性系, 均为福建省1980年选优材料。接穗来源于福建省官庄林场采穗圃。1985年定砧, 1987年嫁接, 1991年开花结实, 1995年进入正常投产期, 1997年底疏伐(强度0.4)。该种子园隔离带为杉木(Cunninghamia lanceolata)种子园、柳杉(Cryptomeria fortunei)天然林、天然阔叶林和人工阔叶林, 宽约300 ~ 500 m。
根据对各无性系种子产量、子代测定、木材密度的历年调查数据, 分别从综合表现高、中、低3个等级中各选出1个有代表性的无性系, 即80、318和335号; 根据3个无性系在种子园内分布的情况(图 1), 分别从园内不同小区的中心部分, 各选择1株作为采种母树, 分别获得了96、94和96株生长正常的后代苗木。将东区范围内所有无性系都作为分析用的候选父本(包括采种母树), 共129个候选父本无性系。每个无性系只取1个单株的种子。

|
图 1 种子园小区分布图 Fig. 1 Illustration of the seed orchard of P.massoniana |
裸子植物的胚乳由未经过受精作用的大配子体发育而来, 带有母本的单倍体信息, 同时符合孟德尔遗传分配规律。多粒种子胚乳的单倍型(haplotype)组合在一起, 就可以代表母树的基因型。一般认为随机选取5粒种子的胚乳进行分析即可满足需要(赖焕林等, 1997), 确定母本基因型的概率可以达到96.9 %。本文对采种母树或候选父本无性系分别选取8粒种子的胚乳来确定其基因型, 准确率可达到99.6 %。子代基因型通过提取子代幼苗的DNA来确定。采用SDS法(尹佟明等, 1997)提取胚乳DNA, 采用CTAB法提取子代幼苗的DNA。
1.3 SSR引物的选择及筛选由于未见关于马尾松的SSR标记应用的报道, 因此本研究选取近缘种、近缘属中应用过的SSR引物作为初筛对象。初筛SSR引物共163对, 分别来自欧洲云杉(Picea abies)、白云杉(Picea glauca)、日本柳杉(Cryptomeria japonica)和松属中的火炬松(Pinus taeda)、北美乔松(Pinus strobus)。最后经复筛得到具有多态性的引物11对(表 1)。
|
|

PCR反应总体系10 μL :20 ng·μL-1的模板DNA 1 μL; 10×缓冲溶液1 μL; 2 mmol· L-1的dNTPs溶液1 μL; 2.5 mmol·L-1的MgCl2溶液1 μL; 10 μmol·L-1的引物各0.25 μL; 5 units·μL-1的AmpliTaq Gold 0.05 μL; 双蒸水5.45 μL。PCR扩增反应采用梯度降温程序:首先94 ℃ 4 min预变性; 然后梯度降温20个循环, 即94 ℃ 30 s, 60 ℃≥50.5 (-0.5 ℃/cyc)30 s, 72 ℃ 30 s; 接着一般性扩增20个循环, 即94 ℃30 s, 50 ℃30 s, 72 ℃30 s; 最后延伸72 ℃5 min; 4 ℃保存。8 %聚丙烯酰胺凝胶垂直电泳, 银染, 拍照。
1.5 数据处理与统计分析采用小源凝胶图像分析系统对所得的银染照片判读。利用软件POPGENE 1.32 (Yeh et al., 2000)计算亲、子代的遗传多样性, 分析的参数有:有效等位基因数(Ne)、观测杂合度(Ho)、期望杂合度(He)、Nei基因多样度指数(h)、固定指数(F)。
将126个无性系和3株采种母树均作为候选父本, 利用软件CERVUS2.0对子代进行父本分析。该软件基于最大似然分配法原理, 要求共显性标记的数据, 可以对双亲都不知道或只知道1个亲本的子代群体作亲本分析; 通过计算机模拟推断可能父本及显著性水平(Marshall et al., 1998)。
2 结果与分析11对SSR引物11个位点的等位基因数由3到20不等, 平均等位基因数为6.818 2个(图 2)。

|
图 2 引物PtTX3116的部分扩增产物 Fig. 2 Part of products amplified by primer PtTX3116 M:标准DNA样品DNA marker; 1~ 18:部分无性系样品Samples of clones. |
从表 2中可以看出, 亲本群体的平均观测杂合度(Ho)为0.343 6, 平均期望杂合度(He)为0.550 6, 平均Nei基因多样度指数(h)为0.543 8;平均固定指数(F)值为0.387 1, 表明亲本群体的杂合体偏少, 纯合体偏多。
|
|
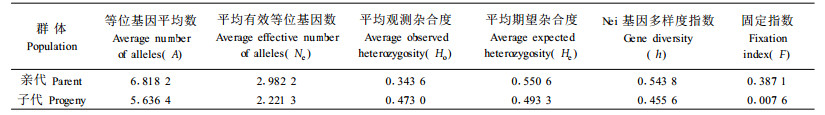
子代群体的平均观测杂合度、平均期望杂合度、平均Nei基因多样度指数分别为0.473 0、0.493 3和0.455 6, 同种子园亲本群体相应的各项参数值差别不显著(表 2); 平均固定指数值为0.007 9, 接近0, 表明3株母树的子代群体是近似于随机交配的产物。80号母树子代的观测杂合度、期望杂合度和Nei基因多样度指数分别为0.443 2、0.411 8和0.409 7, 318号母树子代的为0.472 9、0.486 1和0.483 5, 335号母树子代的为0.502 8、0.476 1和0.473 6, 各项参数值差别不显著。
2.2 父本组成的显著性分析父本分析表明, 除3株母树外, 126个候选亲本中有82个无性系与3株母树产生了后代, 3株母树子代可能的父本组成数量差别不大, 各占种子园无性系数量的30 %左右:80、318和335号母树子代的父本组成数分别为40个、38个和38个。在以往的父本分析研究中, 由于以90 %~ 95 %显著性水平确定父本的比例都不高, 一般都会采用70 %~ 80 %显著性水平对结果作辅助分析(Marshall et al., 1998; 张冬梅等, 2001; 陈小勇, 1999; 陈小勇等, 2000)。本研究中采用95 %和80 % 2种显著性水平。在总共286粒种子中, 有25粒种子可在95 %水平上(P<5 %)确定其父本, 占8.80 %; 107粒种子可在80 %(P<20 %)和95 %水平之间确定父本, 占37.67 %; 在80 %水平上可确定46.47 %种子的父本。其中无性系80号可确定的父本占比例最高, 无性系335号最低(表 3)。本研究显著性分析结果同其他研究者的相近。张冬梅等(2001)在油松(Pinus tabulaeformis)无性系种子的分析中, 80 %显著水平下确定的父本数占55 %; Smouse等(1994)在侏儒百合(Chamaelirium luteum)中取样2 255个后代, 基于排除法分析时只有约1/40的后代数据可以利用; 陈小勇等(2000)对19株青冈(Cyclobalanopsis glauca)实生幼苗的父本分析中, 只有8株能确定唯一亲本, 仅占42.11 %。何田华等(2001)认为, 在排除法研究中, 对大多数亲本分析来说, 所研究的居群中的可能亲本最好不要超过100个, 否则分析中所受的限制将增加。本文共分析了129个候选父本, 确定父本的能力可能受到了一定的影响。
|
|

本研究中, 子代父本组成的成分差别较大, 共6个无性系与所有3株母树都产生了后代; 而与其中2株母树产生后代的有24个无性系; 其他无性系仅与其中1株母树产生后代。无性系起源与亲本可配性无明显关系。没有形成同一起源的几个无性系偏向于与一株母树产生较多子代的情况, 即没有形成固定的交配组合。从表 3可以看出, 对3株母树贡献花粉形成子代比例超过各子代总数5 %的父本数量较少, 分别为6个、6个和2个, 分别占候选父本总数的15.0 %、15.8 %和5.3 %, 但这些父本花粉的总贡献率却分别达到48.96 %、38.30 %和27.08 %; 贡献率低于4 %(大多在1 %~ 2 %)的父本数量较多, 分别为34、28、33个, 分别占候选父本数量的85.0 %、73.7 %和86.8 %, 总贡献率分别达到51.04 %、44.68 %和60.42 %。
有3个候选父本无性系同时为80号和335号母树提供了较多的有效花粉, 对这2个母树子代的贡献率分别达到了25 %和29 %, 其中一个无性系花粉贡献最大, 分别达到了10 %和17 %。张冬梅等(2001)对油松种子园的父本分析获得了相似的结果, 也发现有少数母树花粉来源比较单一, 但大部分母树花粉来源较广。
2.4 种子园外来花粉影响分析本研究中发现了外来花粉影响的现象。在11对SSR引物的电泳结果中共发现候选亲本没有而子代特有的等位基因7个, 分别来自6个SSR位点, 共形成27个后代, 占全部测定后代的9.44 %。从种子园情况看, 种子园周围的隔离带约300 ~ 500 m, 外源花粉从周围的马尾松天然群体流动来的可能性相对较小, 而来源于种子园西区(本文采样的是东区)的可能性较大。尽管东、西区分别在2座山上(图 1), 2区之间也有其他树种相隔, 但它们之间仍有近10 %的花粉交流。考虑到外来花粉(如西区)有同本区花粉的标记基因型相同等情况, 则外来花粉的影响可能高于9.44 %。可见外来花粉污染问题不容忽视, 有效的隔离措施对保证种子的遗传品质是非常必要的。
3 结论与讨论通过对五一林场马尾松无性系种子园亲、子代群体的遗传多样性的分析表明, 该马尾松种子园具有较高的遗传多样性(h =0.543 8, 表 2)。赖焕林等(1997)曾用同工酶对马尾松种子园的遗传多样性进行研究, 获得了相似的结果(h =0.533 5), 研究表明马尾松种子园及其子代的遗传多样性高于天然群体。不过吴若菁(2002)通过醇溶蛋白电泳分析获得了较高的马尾松天然群体的遗传多样性(h =0.552)。本研究的亲本群体的平均Nei基因多样度指数为0.543 8, 和其他松属植物的天然群体相近:北美乔松为0.515(Echt et al., 1996), 油松为0.590, 加勒比松(Pinus caribaea)为0.492, 火炬松为0.513, 日本柳杉为0.666(Moriguchi et al., 2003), 小干松(Pinus contorta)为0.758(Thomas et al., 1999)。以上数据表明, 本研究的亲本遗传多样度并没有因为选择的作用而变小。尽管本研究的种子园子代群体的遗传多样性较亲代群体略低, 但差异并不显著, 而且仅是3株母树的子代群体, 因此可以说明种子园子代群体的遗传多样性不会下降, 同其亲代群体及马尾松天然群体的遗传多样性相近。人们采用同工酶等标记在不同树种研究中也获得了相似的结果(Mitton, 1983; 葛颂, 1989; 王章荣等, 1994; Brown, 1990; 赖焕林, 1995); 也有研究结果表明种子园子代能保持比天然群体较高的遗传多样性(Roberds et al., 1984; 赖焕林等, 1997)。
尽管3株母树在种子园中的位置有所不同, 80号靠种子园东区的中心, 318号靠边缘, 335号介于两者之间(图 1), 但它们子代的父本组成数量几乎没有差别, 分别为40、38和38;它们子代的观测杂合度、期望杂合度和Nei基因多样度指数也没有显著差异。可能是由于马尾松花粉能随风传播很远距离, 种子园中个体所处位置的不同, 并不会对其后代的遗传多样性和父本组成数量造成明显影响。虽然父本花粉贡献率对3株母树有差异(表 3), 但没有明显的规律性, 这可能既与母株所处位置有关, 又与母树本身的遗传特性有关。
本研究中约有5.3 %~ 15.8 %的候选父本贡献了27.08 %~ 48.96 %的花粉, 同时, 有3个父本无性系对80号和335号母树贡献了25 %~ 29 %的有效花粉, 其中一个无性系贡献率分别达到了10 %和17 %。表明种子园内无性系对后代贡献不均衡, 少数无性系花粉占明显的优势。尽管由于子代群体的父本组成较为广泛, 子代的遗传多样性并没有因此而显著变小, 即能维持子代较宽的遗传基础(赖焕林等, 1997), 但仍应给予足够重视, 对这些具明显花粉优势的无性系进行配合力测定, 以期使种子园后代的遗传增益得到保证。
研究的种子园内有一定的外来花粉污染率, 接近10 %, 不过相对其他松属树种的研究结果, 本种子园外花粉的影响较小, 如火炬松36 %~ 60 %、湿地松(P.elliotii)84.5 %、欧洲赤松(P.sylvestris)9 %~ 74 %(赖焕林等, 1997)。究其原因, 与种子园内较多无性系数量及较好的种子园隔离措施有一定关系。
另外, 在父本分析中没有检测到母树的自交现象。由于在研究中未对发育不全的种子及畸形苗木进行父本分析, 这些种子或苗木可能有一部分是自交的结果, 自交衰退现象可能在种子和苗木时期就已出现。当然, 这一推论有待今后对这类种子和苗木进行检测加以验证; 但正常苗木中检测不到自交后代, 使我们可以不用担心用种子园子代造林出现自交衰退的问题。
陈小勇. 1999. 自然植物种群的亲本分析及其在生态学研究中的应用. 生态学杂志, 18(2): 30-35. DOI:10.3321/j.issn:1000-4890.1999.02.008 |
陈小勇, 宋永昌. 2000. 自然定居青冈幼苗的亲本分析. 武汉植物学研究, 18(3): 174-180. DOI:10.3969/j.issn.2095-0837.2000.03.002 |
葛颂. 1989. 用同工酶定量分析林木群体变异和分化的方法. 西南林学院学报, 9(1): 83-91. |
何田华, 葛颂. 2001. 植物种群交配系统、亲本分析以及基因流动研究. 植物生态学报, 25(2): 144-154. DOI:10.3321/j.issn:1005-264X.2001.02.003 |
赖焕林. 1995. 通过种子分析估算交配系统参数的误差. 南京林业大学学报, 19(4): 71-74. |
赖焕林, 王章荣. 1997. 马尾松种子园及其附近人工林的亲子代群体遗传结构分析. 林业科学研究, 10(5): 490-494. DOI:10.3321/j.issn:1001-1498.1997.05.007 |
王章荣, 赖焕林.1994.种子园发展现状与种子园产量和质量的提高∥沈熙环.种子园优质高产技术.北京: 中国林业出版社, 9-13
|
吴若菁. 2002. 天然马尾松群体遗传结构的研究. 林业科学, 38(5): 160-165. DOI:10.3321/j.issn:1001-7488.2002.05.027 |
尹佟明, 黄敏仁, 王明庥, 等. 1997. 利用RAPD标记和单株树大配子体构建马尾松的分子标记连锁图谱. 植物学报, 39(7): 607-612. |
张冬梅, 沈熙环, 何田华. 2001. 利用等位酶对油松无性系种子进行父本分析. 植物生态学报, 25(2): 166-174. DOI:10.3321/j.issn:1005-264X.2001.02.006 |
Brown A H D.1990.Genetic characterization of plant mating systems∥Broen A H D.Plant population genetics, breeding and genetic resources.Sunderland Ma : Sinzuer Associates Inc, 145-162
|
Echt C, Burns R.1999.SSR derived from Pinus taeda ESTs[EB/OL] [1999].http://dendrome.ucdavis.edu.
|
Echt C S, Maymarquardt P, Hseih M, et al. 1996. Characterisation of microsatellite markers in eastern white pine. Genome, 39: 1102-1108. DOI:10.1139/g96-138 |
Elsik C G, Williams C G. 2001. Low-copy micro-satellite recovery from a conifer genome. Theor Appl Genet, 103: 1189-1195. DOI:10.1007/s001220100725 |
Elsik C G, Minihan V T, Hall S E, et al. 2000. Low-copy micro-satellite markers for Pinus taeda L. Genome, 43: 550-555. DOI:10.1139/g00-002 |
Hodgetts R B, Aleksiuk M A, Brown A, et al. 2001. Development of microsatellite markers for white spruce (Picea glauca)and related species. Theor Appl Genet, 102: 1252-1258. DOI:10.1007/s00122-001-0546-0 |
Lexer C, Heinze S, Gerber S, et al. 2000. Microsatellite analysis of maternal half-sib families of Quercus robur, pedunculate oak (Ⅱ):Inferring the number of pollen donors from the offspring. Theor Appl Genet, 100: 858-865. DOI:10.1007/s001220051362 |
Marshall T C, Slate J, Kruuk L E, et al. 1998. Statistical confidence for likelihood-based paternity inference in natural populations. Molecular Ecology, 7: 639-655. DOI:10.1046/j.1365-294x.1998.00374.x |
Mitton J B.1983.Conifers∥Tanksley S D, Orton T J.Isozymes in plant genetics and breeding elsevier.Amsterdam : Science Publishers, Part B, 443-472
|
Moriguchi Y, Iwata H, Ujino-Ihara T, et al. 2003. Development and characterization of microsatellite markers for Cryptomeria japonica D.Don. Theor Appl Genet, 106: 751-758. DOI:10.1007/s00122-002-1149-0 |
Roberds J H, Conkle M T. 1984. Genetic structure in loblolly pine stands:allozyme variation in parents and progeny. For Sci, 30(2): 319-329. |
Smouse P E, Meagher T R. 1994. Genetic analysis of male reproductive contributions in Chamaelirium luteum (L.)Gray (Liliaceae). Genetics, 136: 313-322. |
Thomas B R, Macdonald S E, Hicks M. 1999. Effects of reforestation methods on genetic diversity of lodgepole pine:an assessment using microsatellite and randomly amplified polymorphic DNA markers. Theor Appl Genet, 98: 793-801. DOI:10.1007/s001220051136 |
Yeh F C, Yang R.2000.POPGENE v1.32[CP/OL][2000-12].http://www.ualberta.ca.
|
Zhou Y, Bui T, Auc L D. 2002. Undermethylated DNA as a source of microsatellites from a conifer genome. Genome, 45: 91-99. DOI:10.1139/g01-119 |