 2006, Vol. 42
2006, Vol. 42文章信息
- 张星耀, 赵嘉平, 梁军, 吕全.
- Zhang Xingyao, Zhao Jiaping, Liang Jun, Lü Quan.
- 树木溃疡病菌主要类群系统地位及茶藨子葡萄座腔菌的种内一致性
- Molecular Relationship of genus Botryosphaeria, genus Coniothyrium, genus Dothichiza and the Consistency of Botryosphaeria dothidea
- 林业科学, 2006, 42(11): 63-68.
- Scientia Silvae Sinicae, 2006, 42(11): 63-68.
-
文章历史
- 收稿日期:2005-04-08
-
作者相关文章


树木溃疡病病原真菌类群泛指一些能引起多种树木发生溃疡、烂皮以及枯枝等症状病害的真菌。这些真菌的性质与分类地位十分相近, 据国内外报道有几十种之多, 其中对树木生长造成严重影响的主要是葡萄座腔菌属(Botryosphaeria), 盾壳霉属(Coniothyrium), 疡壳孢属(Dothichiza)以及腐皮壳菌属(Valsa)真菌, 这些真菌对我国人工林, 特别是对杨树人工林造成的损失和危害已有非常多的报道, 被认为是我国现阶段森林重大有害生物之一(张星耀等, 2003)。
关于树木溃疡病病原真菌类群及其所致病害, 国内外众多学者从病原学、病理学、流行学、防治学以及森林生态学等多方面对其进行了大量的研究(Shear et al., 1925; Smith, 1934; 李传道, 1979; 向玉英等, 1979; 1987; 赵仕光, 1993; 1997), 也对关于葡萄座腔菌属的致病机理和遗传分化进行了RFLP、RAPD、ISSR以及ITS序列研究(Jacobs et al., 1998; 张星耀等, 1999; 2000; Smith et al., 2001b; Zhou et al., 2001a; 2001b), 澄清了此类真菌在系统学方面的一些问题。本研究运用RAMs-PCR(random amplification microsatellites-PCR) (Hantula et al., 1996)技术对葡萄座腔菌属与疡壳孢属、盾壳霉属真菌的分类关系, 茶藨子葡萄座腔菌(Botryosphaeria dothidea)的种内遗传一致性, 苹果干腐病病原菌Botryosphaeria berengeriana与苹果轮纹病病原真菌Botryosphaeria berengeriana de Not. f. sp. piricola的相互关系, B. berengeriana与茶藨子葡萄座腔菌的相互关系进行了比较研究, 期望会对溃疡病病原真菌类群的基础生物学研究及树木溃疡病防治提供理论基础。
1 材料与方法 1.1 供试菌株供试菌株包括22个B. dothidea菌株(简记为Bd, 下同), 6个Dothiorella菌株(D), 6个B. berengeriana菌株(Bb), 6个Coniothyrium菌株(Co), 2个Dothichiza菌株(Dp), 其寄主和采集地见表 1。
|
|

以PSA培养基(马铃薯200 g, 葡萄糖20 g, 琼脂20 g, 水1 L)培养供试菌株, 10 d后, 在菌落周缘取直径为3 mm的菌饼8片, 置于150 mL的三角瓶中以PS培养基(马铃薯200 g, 蔗糖20 g, 水1 000 mL), 28 ℃恒温液体静置培养, 10 d后收获菌丝并用灭菌去离子水洗涤3次, -20 ℃保存备用。
1.3 DNA提取DNA提取方法参考张星耀等(1999)的方法。
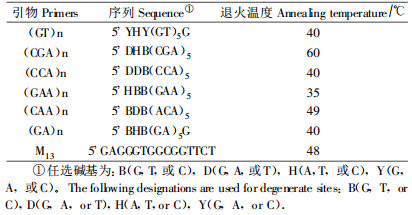
1.4 RAMs-PCR扩增本研究的微卫星引物序列及其退火温度如表 2所示。实验中所用引物由北京九发金贝生物技术有限公司合成, Taq酶购于北京鼎国生物技术有限公司。扩增反应采用25 uL反应体系。包括1×缓冲液(100 mmoL·L-1 Tris HCl, 15 mmol·L-1 MgCl2, 50 mmol·L-1 KCl), 0.4 μmol·L-1的RAMs引物, 0.2 mmol·L-1的dNTPs, 1 U的Taq酶, 模板DNA约10 ng·(25 μL)-1, 反应体系中加入20 μL矿物油以防止反应液蒸发。
|
|
反应条件: 94 ℃变性2 min以后, 94 ℃变性30 s, 退火30 s(各引物退火温度见表 2), 72 ℃延伸45 s为一个循环, 共计35次循环, 72 ℃保持5 min。扩增反应在PE公司DNA Thermal Cycler 480上进行。热启动PCR以防止低温过程中的非特异性扩增。每一引物重复3次, 设不加模板DNA的反应作为阴性对照。
扩增产物在5 %及8 %聚丙烯酰胺凝胶, 1×TBE缓冲液, 120 V电压下电泳, 以鼎国生物工程公司的100 bp DNA Ladder作为分子量标准。硝酸银染色法显色。胶片观察灯下观察, 照相记录。
1.5 RAMs—PCR产物电泳数据的处理将聚丙烯酰胺凝胶电泳图像输入计算机, 利用凝胶分析软件BandScan5.0 (Glyko, Inc.1998)对电泳条带进行识别, 小于10 bp被认为属于同一条带。根据某一条带在样品中出现和不出现, 分别赋值为“1”和“0”, 得到供试菌株的扩增谱带二态数据矩阵。
将扩增谱带二态数据矩阵根据按以下公式计算相关菌株的相似系数I (Nei et al., 1979), I=2Nij (Ni+Nj), 其中, Ni, Nj为2样品的总显示带数, Nij为2样品共有的显示带数, 距离D=1-I。用邻位连接法(neighbor-joining, NJ)对供试菌株进行聚类, 得到树法计算系统树。相似系数的计算和聚类分析均由Ntsys2.02分析软件完成。
2 结果与分析7个引物在供试菌株中扩增产物的分子量大小及各个引物的特征性扩增产物(总计13个) (见表 3)。
|
|

根据扩增结果对供试菌株进行NJ聚类分析, 结果见图 1。

|
图 1 树木溃疡病病原真菌菌株的RAMs-PCR NJ系统进化树 Fig. 1 The NJ phylogeny tree of pathogenic fungi caused trees canker based on RAMs-PCR analysis |
由NJ方法建立的系统树显示:所有的42株供试菌株, 在10.5的水平上可以分为4类, 我们定义为4个RAMs分类群:群Ⅰ包括11株Botryosphaeria dobhidea菌株: A, HX13, B, Y1, Y2, Y3, DB5, C, HK, SP1, YKB; 1株B. berengeriana菌株PGP; 2株Dothiorella gregaria菌株SA, SC。群Ⅱ包括茶藨子葡萄座腔菌菌株PP, Dothiorella菌株SB。群Ⅲ包括6株茶藨子葡萄座腔菌菌株DB2, YKP, YKJ, YKT, TJD, P1, 除TJD为桃树流胶病病原真菌外, 其余均为杨树溃疡病菌; 4株B. berengeriana菌株PB, PT, PGB, PGT; 2株Dothichiza菌株YA, DZT; 6株Coniothyrium菌株SP8, LUK, 4, 5, P10, YKTrm。群Ⅳ由5株茶藨子葡萄座腔菌菌株TJ, P4, Hku, 1, 3; 2株Dothiorella菌株P8, BYK构成。群Ⅲ可以在更近的距离水平上分为2个亚群:亚群1包含了苹果干腐病和轮纹病菌PB, PT, PGB, PGT, 以及溃疡病病原真菌菌株DB2, YKP, YKJ, YKT, TJD, P1;亚群2则由2个Dothichiza菌株和6个Coniothyrium构成。
RAMs-PCR的聚类结果不能将Botryosphaeria、Dothichiza、Coniothyrium属供试菌株区分为3个单一的类群, 系统树显示真菌属间有着比较复杂的亲缘关系。7个茶藨子葡萄座腔菌供试菌株与Coniothyrium, Dothichiza属的供试菌株构成了一个混合的分类群(群Ⅲ)。所有的Coniothyrium菌株则与Dothichiza菌株严格地聚为一类。茶藨子葡萄座腔菌分布于以茶藨子葡萄座腔菌为主构成的3个分类群中, 根据无性阶段确定为Dothiorella的石榴溃疡病菌株SB与苹果轮纹病菌株PP组成一群, 在类群Ⅲ中的茶藨子葡萄座腔菌真菌菌株表现出了与苹果轮纹病及干腐病菌株的较近的亲缘关系, RAMs-PCR的方法并不能将这两种真菌分开。B. dothidea菌株在10.5的水平上形成了4个RAMs分类群(见图 1), 类群间的距离甚至于大于Coniothyrium, Dothichiza属间的遗传距离。
3 结论与讨论 3.1 Botryosphaeria及相关属真菌的RAMs分子标记位点引物M13, (GAA) n, (GT) n、(CGA) n、(AAC) n和(CCA) n均能产生1条以上的类群内共有的特征性微卫星DNA片段, 或者几条微卫星DNA片段共同代表类群的特征, 应用这些引物可以对树木溃疡病原真菌进行快速分子鉴定。
3.2 Coniothyrium与Dothichiza属真菌的分类地位传统分类系统中Botryosphaeria属、Coniothyrium属和Dothichiza属真菌同属腔孢纲(Coelomycetes)、球壳孢科(Sphaeropsidaceae), 以子座、分生孢子器和分生孢子的形态特征来区分。Coniothyrium的分生孢子有色, 小, 圆形或椭圆形, 其他3属分生孢子无色; Dothichiza不具有子座, 分生孢子器为盘状不规则开裂, 分生孢子小于15 μm; Dothiorella具有子座, 而且多个分生孢子器集生在其中, 孢子较大, 圆筒形或卵圆形。张星耀等(1999)通过28SrDNA-RFLP和RAPD技术研究了Coniothyrium属、Botryosphaeria属与Dothichiza属的系统关系, 结果表明Coniothyrium、与Dothichiza属真菌的分类地位较为接近, 而与Botryosphaeria属的亲缘关系相对较远。
本研究中, 在8.0的水平上, 6株Coniothyrium菌株与2株Dothichiza菌株严格聚合在一起, 形成1个复合的RAMs-PCR分类群, 说明Coniothyrium与Dothichiza真菌具有更近的亲缘关系; 除Sp8以外的5个Coniothyrium菌株形成1个小的聚类枝, Dothichiza菌株DZT与YA处于另一聚类枝上, 又说明Coniothyrium与Dothichiza真菌不同。
3.3 杨树溃疡病病原真菌菌株A、B、C及石榴溃疡病菌SB、SB、SC的分子关系菌株A、B、C是杨树溃疡病病原真菌中研究的较为充分的3个菌株。赵仕光(1998)、刘会香等(1997a; 1997b; 1997c)对A、B、C 3菌株的形态特征、生理学特性、同工酶、可溶性蛋白质、致病性及诱导抗性等方面进行了系统研究。结果表明:在形态特征和生理特征方面, 将A、C两菌株归为同一生理型, B菌株属另一生理型。但是诱导作用的大小与生理型之间没有必然联系。根据RAPD-PCR结果表明A、B在分子水平上具有更大的相似性(张星耀等, 2000)。本试验中得到了与后者相似的结论。RAPD分子标记具有较快的进化速率, 因此, 具有一定的鉴别作用, 利用进化更快的微卫星分子标记, 可以看出3菌株之间的相似性下降, 但是来自陕西的A、B菌株仍然聚合在一起, 而与来自于辽宁的菌株C有了较为明显的分化。A、B、C的分化现象显示出了地理起源与B. dothidea种内分化的相关性。电泳图谱容易看出, 在一些特征性的扩增条带上, 部分引物出现了互补现象, 如(CGA) n的1 500, 1 200条带和(CCA) n引物1 450, 1 290条带, 这清晰地说明了微卫星分子标记的共显性特点, 通过对其序列特征的研究, 将有助于对Botryosphaeria真菌相关微卫星位点进化的了解。
菌株SA、SB、SC是近年来严重危害石榴的病原真菌, SA为石榴疮疥病原, SB为石榴果腐病原, SC为石榴干腐病原。RAMs-PCR的结果显示SA和SC之间具有更大的相似性。
3.4 苹果轮纹病菌和苹果干腐病菌的分类地位苹果轮纹病菌和苹果干腐病菌是危害苹果生产的两种病原真菌, 现在将苹果干腐病菌确定为B. berengeriana, 苹果轮纹病菌确定为B. berengeriana de Not. f. sp. piricola (Nose) (中国果树研究所, 1992)。黄春燕等(2001)、张星耀等(1999)利用RAPD-PCR将苹果轮纹病菌与苹果干腐病菌区分开来, RAPD分析不仅可以用于B. berengeriana的鉴定和种内专化型的区分, 而且可用于区分不同寄主、不同地域的菌株。张星耀等(1999)同时发现苹果干腐病菌和杨树溃疡病菌率先聚合在一起, 表明了两病害病原菌的一定相似性。
本研究的结果未能将苹果干腐病菌与苹果轮纹病菌完全分开, 其中干腐病菌PGP, PGT, 轮纹病菌PB, PT各自聚为一类, 表明了这2类真菌的不同; 但是PGP和PP却聚合在一起。RAMs-PCR的结果显示苹果干腐病菌与苹果轮纹病菌的更多分化现象, 发现来自北京的PB, PP, PGP有着较好的相似性, 而2株山东的菌株与来自北京的PGB有着较好的相关性。
3.5 茶藨子葡萄座腔菌的种内一致性近年来许多研究认为葡萄座腔菌属(Botryosphaeria)不是一个单系类群(Jacobs et al., 1998; Smith et al., 2001a; Zhou et al., 2001a), 其中形态学上确定的茶藨子葡萄座腔菌和B. ribis之间有很大的不同。Ma等(2002)用MP-PCR及ITS序列分析均将86个被认为是茶藨子葡萄座腔菌的菌株, 分为清晰的3个组。张星耀等(2000)的RAPD结果也显示出了茶藨子葡萄座腔菌内部的不一致性, 部分菌株构成了一个纯正的Bd分类群, 另外一部分则与B. berengeriana构成了复合类群。
本研究表明:在我国引起树木溃疡病的病原真菌B. dothidea菌株之间具有较大的遗传分化, 在10.0的水平上, 28个供试菌株被分为2大类, 其中以A, B为代表的一类几乎全由B. dothidea构成, 而另外一类则具有更为复杂的结构, 与Coniothyrium与Dothichiza以及茶藨子葡萄座腔菌的大多数菌株更为相近, 相互之间有着较远的进化距离。在9.0的水平上, B. dothidea则可以分为4个分类群。
在本研究建立的NJ系统树中, B. dothidea类群表现出了内部遗传分化的明显不一致性, 与国外的研究相一致。关于树木溃疡病的病原, 国外有些研究者认为主要是由B. ribis与茶藨子葡萄座腔菌等构成, 即认为B. ribis与茶藨子葡萄座腔菌分别代表 2个不同的真菌类群, 而国内一般认为B. ribis是茶藨子葡萄座腔菌的同物异名。本研究中根据后一种观点将有关的树木溃疡病菌鉴定为茶藨子葡萄座腔菌, 但是试验结果却清晰地揭示出供试菌株的遗传分化。因此, 对于RAMs分析的结果, 如果考虑到部分B. ribis菌株的存在, 则本研究结果与国外完全一致。本研究的结果显示出茶藨子葡萄座腔菌内部的不一致性, 形态学上认定的茶藨子葡萄座腔菌菌株应该是2个到4个种组成的多元类群, B. ribis不是茶藨子葡萄座腔菌的同物异名, RAMs-PCR可以看出二者之间的遗传差异, 因而应该认为是2个不同的生物学种。在与国外的菌株进行比较研究之后, 可以最终验证这个结论的正确性。
黄春燕, 刘开启. 2001. 苹果轮纹病及相关病害病原菌的RAPD分析. 植物病理学报, 31(2): 164-169. DOI:10.3321/j.issn:0412-0914.2001.02.012 |
李传道. 1979. 树木的溃疡病. 南京林学院学报, 1(2): 22-24. |
刘会香, 胡景江, 朱玮, 等. 1997a. 杨树溃疡病菌3菌系脂酶同工酶, 可溶性蛋白及致病性研究. 西北林学院学报, 12(3): 24-28. |
刘会香, 景耀, 赵仕光, 等. 1997b. 杨树溃疡病菌3菌系形态和生理特性的研究. 西北林学院学报, 12(3): 18-23. |
刘会香, 朱玮, 胡景江, 等. 1997c. 杨树溃疡病菌3菌系及诱导物对寄主两种酶活性的影响. 西北林学院学报, 12(3): 29-34. |
向玉英, 花晓梅. 1979. 杨树水泡性溃疡病的病原鉴定. 微生物学报, 19(1): 57-63. |
向玉英. 1987. 杨树病害及其防治. 北京: 中国林业出版社, 55-60.
|
张星耀, 骆有庆. 2003. 中国森林重大生物灾害. 北京: 中国林业出版社, 93-139; 372-385.
|
张星耀, 赵仕光, 吕全, 等. 2000. 树木溃疡病病原真菌类群分子遗传多样性研究Ⅱ—Botryosphaeria属28SrDNA PCR RFLP和RAPD解析. 林业科学, 36(2): 76-81. |
张星耀, 赵仕光, 朴春根, 等. 1999. 树木溃疡病病原真菌类群分子遗传多样性研究Ⅰ—小穴壳菌属, 疡壳孢属, 壳囊孢属, 盾壳霉属分类地位的分子证明. 林业科学, 35(3): 34-40. DOI:10.3321/j.issn:1001-7488.1999.03.007 |
赵仕光.1998.杨树溃疡病寄主组织病理理学和病菌分子生物学研究.东北林业大学博士论文 http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y300778
|
赵仕光, 景耀, 王颍, 等. 1997. 杨树溃疡病测报技术的研究. 西北林学院学报, 12(3): 45-51. |
赵仕光, 景耀, 杨俊秀. 1993. 杨树树皮内过氧化物酶和多酚氧化酶活性与抗溃疡病的关系. 西北林学院学报, 8(3): 11-17. |
中国果树研究所. 1996. 中国果树病虫志. 北京: 中国农业出版社.
|
Hantula J, Dusabenyagasani M, Hamelin C. 1996. Random amplified microsatellite (RAMS)-a novel method for characterizing genetic variation within fungi. Eur.J For Path, 26: 159-166. DOI:10.1111/j.1439-0329.1996.tb00720.x |
Jacobs K A, Rehner S A. 1998. Comparison of cultureal and morphological characters and its sequences in anamorphs of Botryosphaeria and related taxa. Mycologia, 90(4): 601-610. DOI:10.1080/00275514.1998.12026949 |
Ma Zhonghua, Michailides T J. 2002. Character ization of Botryosphaeria dothides isolates collected from pistachio and other plant hosts in California. Pohytopathology, 92: 519-526. DOI:10.1094/PHYTO.2002.92.5.519 |
Nei M, Li W H. 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc Natl Acad Sci, 76: 5269-5273. DOI:10.1073/pnas.76.10.5269 |
Shear C L, Stevens N E, Wilcox M S. 1925. Botryosphaeria and Physalospora in the eastern United States. Mycologia, 17: 98-107. DOI:10.1080/00275514.1925.12020461 |
Smith D R, Stanosz G R. 2001a. M olecular and morphological differentiation of Botryosphaeria dothidea (anamorph Fusiciccum aesculi)from some other fungi with fisicoccum anamorphs. Mycologia, 93(3): 505-515. DOI:10.1080/00275514.2001.12063183 |
Smith H, Crous P W, Wingfield M J, et al. 2001b. Botryosphaeria eucalytorum ap.Nov, a new species in the B. dothidea complex on Eucalytus in South Africa. Mycologia, 93(2): 277-285. DOI:10.1080/00275514.2001.12063159 |
Smith C O. 1934. Inoculations showing the wide host range of Botryosphaeria ribis. Jour Agric Res, 49: 467-476. |
Zhou Shiguo, Smith D R, Stanosz G R, et al. 2001a. Diffention of Botryosphaeria species and related anamorphic fungi using Inter-simple or short sequence repeat (ISSR)fingerprinting. Mycological Research, 105(8): 919-926. DOI:10.1016/S0953-7562(08)61947-4 |
Zhou Shiguo, Stanosz G R. 2001b. Relationships among Botryosphaeria species and associated anamorphic fungi in ferred from the analysis of ITS and 5.8S rDNA sequences. Mycologia, 93(3): 516-527. DOI:10.1080/00275514.2001.12063184 |