 2006, Vol. 42
2006, Vol. 42文章信息
- 关德新, 吴家兵, 金昌杰, 韩士杰, 张弥, 施婷婷.
- Guan Dexin, Wu Jiabing, Jin Changjie, Han Shijie, Zhang Mi, Shi Tingting.
- 长白山红松针阔混交林CO2通量的日变化与季节变化
- Diurnal and Seasonal Variation of CO2 Flux Above the Korean Pine and Broad-Leaved Mixed Forest in Changbai Mountain
- 林业科学, 2006, 42(10): 123-128.
- Scientia Silvae Sinicae, 2006, 42(10): 123-128.
-
文章历史
- 收稿日期:2005-07-08
-
作者相关文章


森林是陆地上面积最大的生态系统, 它通过光合作用吸收大量的CO2并将其长期地储存于生态系统中, 森林生态系统CO2通量的研究备受关注(方精云等, 2001; 于贵瑞等, 2001; 吴家兵等, 2003)。长白山红松(Pinus koraiensis)针阔混交林是东北亚典型的温带植被类型, 因具有较大的面积和较高的净初级生产力而成为森林碳循环研究的重要对象, 被列入全球变化研究的中国东北样带(周广胜等, 2003), 但源库关系、源汇强度等关键数据还不清楚, 特别是弄清昼夜循环和季节更替导致的CO2通量日季变化, 是分析其碳收支机理的基础, 由于观测方法的历史局限, 生态系统尺度的相关研究很难见到。以往有关研究主要观测森林生态系统的某个组分的CO2通量, 如森林土壤(蒋高明等, 1997; 刘绍辉等, 1998; 易志刚等, 2003)、植物体或叶片(李意德等, 1997; 曾小平等, 2000)、树干(Wang et al., 2003)等, 或采用间接计算(方精云, 1999)及大气边界层动力学方法(陈步峰等, 2001)进行生态系统尺度的CO2通量计算, 但观测时间较短。20世纪90年代后期涡动相关技术发展成熟, 开始应用于生态系统尺度的长期CO2通量监测, 2001年全球通量网(FLUXNET)统计的140通量监测站中森林站就有80个(Baldocchi et al., 2001), 涡动相关技术成了多个生物圈研究计划的基本手段。中国通量网(ChinaFLUX)于2002年建成, 包括5个森林监测站, 通过这些监测站, 基本了解了森林生态系统的CO2通量概况(关德新等, 2004; 吴家兵等, 2004; 王秋凤等, 2004; 刘允芬等, 2004; 李正泉等, 2004), 但CO2通量日变化和季节变化规律的研究结果极少。本文在获取周年涡动相关CO2通量数据的基础上, 分析长白山红松针阔混交林生态系统CO2通量的日变化和季节变化特征, 为森林碳收支的研究提供基础。另外, 本研究对象的优势树龄在200年左右, 这在国际碳通量研究中是极少见的, 所以本研究结果可为国际上有关林龄与碳通量关系的研究热点提供重要依据。
1 材料与方法 1.1 试验地概况研究在中国科学院长白山森林生态系统定位站1号标准地红松针阔混交林内进行(42°24′N, 128°6′E), 海拔738 m。根据1982—2003年气候数据, 年均气温3.6 ℃, 年均降雨量695 mm。无霜期140 d左右, 日照时数1 800~2 300 h。土壤为山地暗棕色森林土。观测场附近下垫面地势平坦, 林型为成熟原始林, 主乔木为红松、椴树(Tilia amurensis)、蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandshurica)、色木槭(Acer mono)。林分为复层结构, 下木覆盖度40%, 林分平均株高26 m, 立木株数约560株·hm-2, 优势树龄在200年左右。
1.2 观测方法观测林地建有高62 m的微气象观测塔, 涡动相关系统的探头安装在40 m高度的伸展臂上, 由三维超声风温仪(CSAT3, Campbell, USA)和开路CO2/H2O分析仪(Li-7500, LiCor Inc., USA)组成, 原始采样频率为10 Hz, 数据传输给数据采集器(CR5000, Campbell Scientific, USA)进行存储, 同时进行30 min的通量计算并存储。另外还安装了常规气象观测系统, 包括7层风速仪(A100R, Vector Instruments, Denbighshire, UK)、大气温度和湿度(HMP45C, Vaisala, Helsinki, Finland), 高度分别为2.5、8、22、26、32、50、60 m, 光合有效辐射传感器(LI-190Sb, LiCor Inc., USA)设于距地面32 m处, 地温探头(105T, Campbell, USA)设置深度为0、5、10、20、50、100 cm, 土壤水分TDR探头(CS616-L, Compbell, USA)设置深度为5、20、50 cm, 这些要素的原始采样频率为2 Hz, 通过数据采集器(CR23X and CR10X, Campbell Scientific, USA)采集并按30 min计算平均值进行存储。本文取2003年全年的数据进行分析。
用便携式叶面积仪(Li-2000, LiCor Inc., USA)观测叶面积, 在林内铁塔附近约100 m长的样线上设10个定位点采集原始数据, 采样时光感应探头高度为1 m, 对照点设在塔上32 m高度, 重复2次游动观测, 得到叶面积, 生长季每5~10 d观测1次, 叶生长迅速的春季和落叶期增加观测次数。观测值代表的是高1m以上的叶、干和枝的面积系数PAI (plant area index)。
1.3 涡动相关数据的计算与处理根据三维超声风温仪和开路CO2/H2O分析仪观测资料, 以30 min为平均长度, 采用涡动相关方法实时计算40 m高度的CO2通量FC, 计算公式为
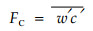
|
(1) |
式中:w'和c'表示重点风速和CO2浓度的脉动值, 上横线表示协方差, 计算过程中还包含了密度订正(Webb et al., 1980)。为了检验探头姿态, 采用三维旋转(3D rotation)和平面旋转(planar-fit rotation)方法进行了坐标订正(Wilczak et al., 2001; 吴家兵等, 2004), 通量结果偏差很小, 年总量变化为0.5%~3.2%, 可以忽略不计。
探头高度以下的大气CO2储存项FS利用分析仪测定的CO2浓度进行计算(Hollinger et al., 1994; Pilegaard et al., 2001; Knohl et al., 2003), 则净交换量(net exchange estimate, NEE) (以下仍称为通量)为NEE=FC+FS, 通量为负值表示森林吸收大气CO2, 正值表示向大气释放CO2。
为了控制数据质量, 从时间系列中剔除方差大于3的数据, 并根据超声风温仪观测的摩擦速度剔除弱湍流条件(摩擦速度小于0.2 m·s-1)的数据, 临界摩擦速度0.2 m·s-1根据夏季(6—8月)夜间的CO2通量与摩擦速度的经验关系确定(赵晓松等, 2006; Guan et al., 2006), 这与其他地区森林的研究结果相似(Goulden et al., 1996; Lindroth et al., 1998)。
全年有效的观测数据占86.9%, 降水期间探头探测路径受阻或停电和仪器维护等原因造成的缺测利用以下方法插补:1)小于3 h的短时间缺测, 用线性内插方法插补; 2)大于3 h的缺测, 按10 d分段的阶段数据规律进行插补, 白天和夜间采用不同的方法, 白天净生态系统CO2交换量(NEE)与光合有效辐射(photosynthetically active radation, PAR)的关系用Michaelis-Menten方程拟合(Morén et al, 2000)
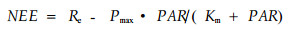
|
(2) |
式中:Pmax为最大光合速率, Re为生态系统呼吸, Km为Michaelis-Menten方程常数, 都与生态系统和季节有关(关德新等, 2004; Guan et al., 2006)。
夜间NEE与土壤温度的关系用指数方程拟合(Lee et al., 1999) :
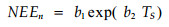
|
(3) |
式中:NEEn为夜间净生态系统CO2交换量, 即生态系统呼吸, Ts为5 cm土壤温度, b1、b2是常数, b1=0.035 6, b2 =0.129 6, R2=0.465, 回归方程达极显著水平(Guan et al., 2006)。
2 结果与分析 2.1 晴天和多云天日变化的比较图 1是生长季夏天典型的多云天(6月18日, 图 1a)和晴天(6月19日, 图 1b)生态系统CO2通量NEE的日变化情况, 图中还绘出了储存项Fs、光合有效辐射PAR、空气饱和差(vapor pressure deficit, VPP)的日变化(18日早3~6时因降雨NEE和Fs缺测)。可见:1)生长季NEE存在明显的日变化, 日出后随光照增加生态系统由CO2释放转为吸收, 晴天条件下吸收强度迅速增大, 在9时左右达到最大(图 1b); 多云天吸收强度与PAR正相关(图 1a); 在日落前由吸收转为释放, 夜间保持释放。2) Fs在日出后由正值转为负值, 一般在早晨7~8时达最小, 白天多为负值, 即探头高度以下的大气CO2浓度由于植被的光合吸收而逐渐下降, Fs在午后比NEE提早转为正值, 夜间保持正值, 即探头高度以下的大气CO2浓度由于生态系统的呼吸作用而逐渐上升。3) CO2的吸收不随PAR的增加而一直增加, 晴天最大的通量出现在上午8~10时, 而不出现在PAR最大的中午, 这主要是由空气饱和差VPD所制约的, 晴天中午高VPD迫使叶片的光合作用下降(Leuning, 1995; Guan et al., 2006), 如图 1b 19日上午8:30 CO2吸收强度最大, 对应的PAR不是全天最大值, 9:00~14:30的PAR均高于此值, 但CO2吸收强度却在逐渐下降。VPD较低时对叶片光合作用的限制作用较小, 吸收强度与PAR的正相关关系较明显(图 1a)。

|
图 1 多云天(a)和晴天(b)生态系统CO2通量(NEE)、储存项(Fs)、光合有效辐射(PAR)和饱和差(VPD)的日变化 Fig. 1 Diurnal variations of CO2 flux (NEE), storage term (Fs), photosynthetically active radiation (PAR) and vapor pressure deficit (VPD) in cloudy (a) and clear day (b) |
图 2为NEE月平均的日变化, 除了寒冷的冬季外, 其他季节CO2通量都具有上述的日变化特征, 即白天吸收、夜间释放CO2, 但强度随季节变化, 大致可分为3个水平, 6—8月白天吸收和夜间释放CO2强度均较大, 月平均最大强度分别达-0.8和0.4 mgC·m-2s-1, 5月和9月次之, 白天和夜间的平均最大强度达-0.4和0.2 mgC·m-2s-1, 其他月份的通量较小, 平均值为-0.1~0.1 mgC·m-2s-1, 其中温度最低的12月至次年2月的通量最小。

|
图 2 CO2通量月平均的日变化 Fig. 2 Monthly averaged diurnal variation of CO2 flux |
CO2通量日总量定义为0点开始累计的24 h通量总和, 2003年日总量NEEd动态变化如图 3所示, 1—3月温度很低, 植物基本没有光合作用, 森林生态系统有微弱的呼吸(平均值0.5 gC·m-2d-1)。随着温度升高, 植物开始萌动, 针叶树种白天可以进行光合作用, 4月中旬以后, NEEd有负值出现, 5月中旬到9月上旬是植被生长旺季, NEEd几乎都是负值, 其中6月上、中旬CO2通量较大, 可超过-6 gC·m-2d-1, 主要原因是阴雨天少、光照充足。7、8月由于阴雨天增多, 通量变小, 偶尔出现净释放(发生在光照很弱、而温度又相对较高的天气)。9月叶片逐渐老化, 光合吸收迅速下降, 10月以后NEEd保持正值。

|
图 3 CO2通量日总量(NEEd)的季节变化 Fig. 3 Seasonal variation of daily CO2 flux (NEEd) |
按月累计的结果, 6月生态系统的净碳吸收最大, 为-110 gC·m-2, 7、8、5月次之, 分别为-81、-60和-41 gC·m-2, 4月的碳交换近似平衡(-1 gC·m-2), 9月为较小负值(-12 gC·m-2), 10和11月正值相对较大(26和23 gC·m-2), 12月和1—3月变化于15~20 gC·m-2之间。2003年该生态系统净碳吸收量为-184 gC·m-2, (相当于-1.84 tC·hm-2), 这与加拿大安大略省南部不足百年林龄落叶混交林1996年的观测结果(190 gC·m-2) (Lee et al., 1999)相近。丹麦80年林龄的榉木林2年的观测结果是232和144 gC·m-2 (Pilegaard et al., 2001), 2年的平均值与本文的结果接近。
2.4 CO2通量日、季变化主要控制因子从气候特点看, 该生态系统降水较充沛, 水分不是影响CO2交换的限制性因子。在年时间尺度上, 叶面积变化是影响生态系统光合吸收CO2的重要原因, 图 4是月平均叶面积指数(plant area index, PAI)随月平均气温Ta的变化, 可用一元三次回归方程拟合
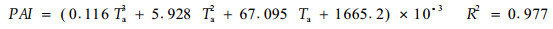
|
(4) |

|
图 4 月平均叶面积指数(PAI)与月平均气温(Ta)的关系 Fig. 4 Relationship between monthly mean plant area index (PAI) and monthly mean air temperature (Ta) |
对回归方程进行显著性检验, p < 0.001, 达极显著水平, 可见2者有密切的相关关系。
由于温度也是生态系统呼吸的主要控制因子(公式3), 所以生态系统CO2通量季节变化主要受温度的制约, 月净交换量NEEm与月平均气温Ta的关系如图 5所示, 可用一元三次回归方程拟合
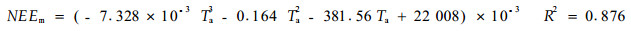
|
(5) |

|
图 5 CO2月通量(NEEm)与月平均气温(Ta)的关系 Fig. 5 Relationship between monthly CO2 flux (NEEm) and monthly mean air temperature (Ta) |
对回归方程进行显著性检验, p < 0.001, 达极显著水平, 可见温度对NEE季节变化制约作用是显著的。
日变化是叠加在季节变化之上的短周期波动, 在其他因素(叶面积、温度、水分供应等)相对稳定时, 生态系统的呼吸也相对稳定, CO2通量日变化主要受光合作用的制约, 所以光合有效辐射PAR是主要影响因子, 图 6显示了7月白天NEE随PAR的变化, 可见2者具有密切的关系, 通常用公式(2)表达。由于温度、叶面积等环境因子的季节变化, 式(2)中的系数也随之变化, 研究结果表明(关德新等, 2004), 该生态系统的Pmax和Re在7月最大, 6、8月次之, 5、9月再次, 这与温度、叶面积的变化特点是一致的, 5—9月回归方程(2)的相关系数为0.548~0.616 (均达达极显著水平)。另外, VPD也是通过影响Pmax和Re起作用的, 这2个系数随VPD的上升而减小, 并与季节有关(关德新等, 2004)。赵晓松等(2006)利用相同观测数据(30 min平均值)进行了生长季各月NEE与环境因子的偏相关分析, 结果表明:如果固定温度和VPD的影响, NEE与PAR的偏相关系数在0.5以上, 其中4—8月在0.6以上; 如果固定温度和PAR的影响, VPD与NEE的4—6月偏相关系数约为0.4, 7—9月为0.2~0.3, 生长季各月温度与NEE的偏相关系数(0.1~0.4)都比PAR与NEE的要小。这些结果表明, CO2通量日变化主要受PAR的制约, VPD也有一定的影响, 而受温度的影响较小。

|
图 6 7月的白天CO2通量(NEE)与光合有效辐射(PAR)的关系 Fig. 6 Relationship between daytime CO2 flux (NEE) and photosynthetically active radiation (PAR) in July |
红松针阔混交林CO2通量随着气象条件的变化呈现明显的日变化和季节变化, 在生长季白天为明显的碳汇, 夜间为碳源, 晴天最大的吸收强度发生在上午9时左右, 而多云天和阴天吸收强度与光照强度同步, 夜间森林生态系统的呼吸强度随环境温度的升高而增大。季节变化特点是生长季生态系统表现为明显的碳吸收, 6—8月吸收量较大, 植物休眠的冬季生态系统表现为弱的碳释放, 生长季与休眠期的过渡阶段碳收支近似平衡。在当地气候条件下, 温度是限制叶面积生长的主要环境因子(如回归方程4), 也是影响呼吸的主要环境因素(如公式2), 所以生态系统CO2通量在年尺度上的季节变化主要受温度的制约(如回归方程5)。而日变化主要受光合有效辐射和饱和差的影响, 温度的影响较小。
森林生态系统CO2通量的日变化和季节变化是系统内各个组分复杂的生理过程和物理过程综合作用的结果, 是森林碳收支机制分析的重要内容。由于生态系统类型、组分和生理、生态过程的复杂性, 不同森林生态系统CO2通量的大小和动态变化也是复杂的, 驱动因子也不尽相同, 特别是不同气候带的森林生态系统, 其制约因子和影响规律可能有较大差别。要弄清森林生态系统碳收支机理, 有必要将生态系统尺度的涡动相关观测与系统中各组分CO2通量观测相结合, 在同步观测的基础上, 针对当地的气候、植被、土壤特点, 探讨制约CO2通量的主要因子和影响规律, 这也是国际上森林碳循环研究的主要趋势之一。
值得注意的是, 各种研究结论都是建立在观测结果之上或需要观测结果来验证的, 所以观测技术和方法显得十分重要。各种观测方法都有优势和不足, 箱式-浓度分析法可以灵活地选择生态系统的不同组分(叶片、土壤、枝干等), 在小尺度上进行观测, 且仪器设备价格较低, 但必须干扰环境, 还存在尺度上推计算中的不确定性, 以及自动观测难度大等不足; 涡动相关方法能够直接观测生态系统尺度的通量, 不干扰环境, 时间分辨率和自动观测程度高, 但诸如地势不平、弱湍流影响、平流干扰等技术问题的存在, 使得该方法还有一定的误差。不同方法之间进行弥补和比较, 是该研究领域今后的重要内容之一。
陈步峰, 林明献, 李意德, 等. 2001. 海南尖峰岭热带山地雨林近冠层CO2及其通量特征研究. 生态学报, 21(12): 2166-2172. DOI:10.3321/j.issn:1000-0933.2001.12.029 |
方精云, 朴世龙, 赵淑清. 2001. CO2失汇与北半球中高纬度陆地生态系统的碳汇. 植物生态学报, 26: 594-602. DOI:10.3321/j.issn:1005-264X.2001.05.014 |
方精云. 1999. 森林群落呼吸量的研究方法及其应用的探讨. 植物学报, 41(1): 88-94. |
关德新, 吴家兵, 于贵瑞, 等. 2004. 气象条件对长白山阔叶红松林CO2通量的影响. 中国科学:D辑, 34(增刊Ⅱ): 103-108. |
蒋高明, 黄银晓. 1997. 北京山区辽东栎林土壤释放CO2的模拟实验研究. 生态学报, 17(5): 476-482. |
李意德, 吴仲民, 曾庆波, 等. 1997. 尖峰岭热带山地雨林群落呼吸量初步测定. 林业科学研究, 10(4): 348-355. DOI:10.3321/j.issn:1001-1498.1997.04.002 |
李正泉, 于贵瑞, 温学发, 等. 2004. 中国通量观测网络(ChinaFLUX)能量平衡闭合状态的评价. 中国科学:D辑, 34(增刊Ⅱ): 46-56. |
刘绍辉, 方精云. 1998. 北京山地温带森林的土壤呼吸. 植物生态学报, 22(2): 119-126. |
刘允芬, 宋霞, 孙晓敏, 等. 2004. 千烟洲人工针叶林CO2通量季节变化及其环境因子的影响. 中国科学:D辑, 34(增刊Ⅱ): 109-117. |
王秋凤, 牛栋, 于贵瑞, 等. 2004. 长白山森林生态系统CO2和水热通量的模拟研究. 中国科学:D辑, 34(增刊Ⅱ): 131-140. |
吴家兵, 张玉书, 关德新. 2003. 森林生态系统CO2通量研究方法与进展. 东北林业大学学报, 31(6): 49-51. DOI:10.3969/j.issn.1000-5382.2003.06.017 |
吴家兵, 关德新, 于贵瑞, 等. 2004. 长白山阔叶林红松林CO2交换的涡动通量修订. 中国科学:D辑, 4(增刊Ⅱ): 95-102. |
易志刚, 蚁伟民, 周国逸, 等. 2003. 鼎湖山三种主要植被类型土壤碳释放研究. 生态学报, 23(8): 1673-1678. DOI:10.3321/j.issn:1000-0933.2003.08.027 |
于贵瑞, 牛栋, 王秋凤. 2001. 《联合国气候变化框架公约》谈判中的焦点问题. 资源科学, 23(6): 10-16. DOI:10.3321/j.issn:1007-7588.2001.06.003 |
赵晓松, 关德新, 吴家兵, 等. 2006. 长白山阔叶林红松林CO2通量与温度的关系. 生态学报, 26(4): 16-23. |
曾小平, 彭少麟, 赵平. 2000. 广东南亚热带马占相思林呼吸量的测定. 植物生态学报, 24(4): 420-424. DOI:10.3321/j.issn:1005-264X.2000.04.006 |
周广胜, 王玉辉, 许振柱, 等. 2003. 东北样带碳循环研究进展. 自然科学进展, 13(9): 917-922. DOI:10.3321/j.issn:1002-008X.2003.09.004 |
Baldocchi D, Falge E, Gu L H, et al. 2001. FLUXNET :a new tool to study the temporal and spatial variability of ecosystem -scale carbon dioxide, water vapor, and energy flux densities. Bulletin of American Meteorological Society, 82(11): 2415-2434. DOI:10.1175/1520-0477(2001)082<2415:FANTTS>2.3.CO;2 |
Goulden M L, Munger J W, Fan S M, et al. 1996. Exchange of carbon dioxide by a deciduous forest :response to interannual climate variability. Science, 271: 1576-1578. DOI:10.1126/science.271.5255.1576 |
Guan Dexin, Wu Jiabing, Zhao Xiaosong, et al. 2006. CO2 fluxes over an old, temperate mixed forest in North-eastern China. Agricultural and Forest Meteorology, 137(3-4): 130-149. |
Hollinger D Y, Kelliher F M, Byers J N, et al. 1994. Carbon dioxide exchange between an undisturbed old-growth temperate forest and the atmosphere. Ecology, 75(1): 143-150. |
Knohl A, Schulze A, Kolle O, et al. 2003. Large carbon uptake by an unmanaged 250-year-old deciduous forest in Central Germany. Agricultural and Forest Meteorology, 95: 115-168. |
Lee X, Fuentes J D, Staebler R M, et al. 1999. Long-term observation of the atmospheric exchange of CO2 with a temperate deciduous forest in southern Ontario, Canada. Journal of Geophysical Research, 104: 15975-15984. DOI:10.1029/1999JD900227 |
Leuning R. 1995. A critical appraisal of a combined stomatalphotosynthesis model for C3 plants. Plant, Cell and Environmen, 18: 339-355. DOI:10.1111/j.1365-3040.1995.tb00370.x |
Lindroth A A, Grelle A, Moren A S. 1998. Long-term measurements of boreal forest carbon balance reveal large temperature sensitivity. Global Change Biology, 4: 443-450. DOI:10.1046/j.1365-2486.1998.00165.x |
Morén A S, Lindroth A. 2000. CO2 exchange at the floor of a boreal forest. Agricultural and Forest M eteorology, 101: 1-4. DOI:10.1016/S0168-1923(99)00160-4 |
Pilegaard K, Hummelshø j P, Jensen N O, et al. 2001. Two years of continuous CO2 eddy-flux measurements over a Danish beech forest. Agricultural and Forest Meteorology, 107: 29-41. DOI:10.1016/S0168-1923(00)00227-6 |
Wang Wenjie, Yang Fengjian, Zu Yuangang. 2003. Stem respiration of a larch plantation in northeast China. Acta Botanica Sinica, 45(12): 1387-1397. |
Webb E K, Pearman G I, Leuning R. 1980. Correction of flux measurements for density effects due to heat and water vapour transfer. Quarterly Journal of the Royal Meteorological Society, 106: 85-100. DOI:10.1002/qj.49710644707 |
Wilczak J M, Oncley S P, Stage S A. 2001. Sonic anemometer tilt correction algorithms. Boundary-Layer Meteorology, 99(1): 127-150. DOI:10.1023/A:1018966204465 |