 2006, Vol. 42
2006, Vol. 42文章信息
- 付增娟, 张川红, 郑勇奇, 王志和, 段福文.
- Fu Zengjuan, Zhang Chuanhong, Zheng Yongqi, Wang Zhihe, Duan Fuwen.
- 黑荆和银荆的繁殖扩散与入侵潜力
- Invasive Potential of Two Introduced Tree Species:Acacia mearnsii and Acacia dealbata
- 林业科学, 2006, 42(10): 48-53.
- Scientia Silvae Sinicae, 2006, 42(10): 48-53.
-
文章历史
- 收稿日期:2005-01-13
-
作者相关文章


2. 云南省楚雄彝族自治州林业科学研究所 楚雄 675000
2. Chuxiong Institute of Forestry, Yunnan Province Chuxiong 675000
外来种入侵已经对生态环境和经济发展产生巨大影响(Vitousek et al., 1997; Chapin et al., 2000), 成为生物多样性丧失的两大主要原因之一, 仅次于生境破坏(Wilcove et al., 1998)。决定外来植物入侵的自身因素包括繁殖与传播特性、生态幅和遗传结构(徐汝梅等, 2003; 徐承远等, 2001)。其中, 外来植物的繁殖能力大小对能否形成入侵具有重要影响, 一般认为, 植物的繁殖能力与其入侵能力呈正相关(Barret, 1983)。通常, 入侵性植物都具有性和无性两种繁殖方式, 且表现出种子产量高、幼苗期短、种子发芽率高、幼苗生长快等特点。一个成功的入侵种, 能造成显著或严重的经济或生态影响, 种群密度大和空间分布广是必备的条件, 而物种的扩散又有赖于它的繁殖能力、扩散方式和扩散能力(徐汝梅, 2003)。对于尚未造成入侵灾害的外来树种, 研究其繁殖能力和扩散潜力显得更为重要(Poschlod et al., 2000), 它有助于分析外来种的入侵潜力, 预测潜在入侵风险, 以便制定合适的入侵防控策略。
黑荆(Acacia mearnsii)和银荆(Acacia dealbata)都原产于澳大利亚, 天然分布于新南威尔士、维多利亚、南澳大利亚州及塔斯马尼亚岛。19世纪50年代初引入我国, 由于用途广泛、生长快、能固氮、可以改良土壤, 已经在我国南方各省大面积栽培(潘志刚等, 1994)。这两个树种在国外都有很长的入侵历史。黑荆已经在新西兰、夏威夷、南非和津巴布韦形成入侵(Smith, 1985; Quentin et al., 1995; Maitre et al., 2002; Nyoka, 2003), 被列为世界100种恶性外来入侵物种之一(Lowe et al., 2000); 银荆在西班牙、南非和津巴布韦形成入侵(Hendersong, 1984; Maitre et al., 2002; Nyoka, 2003), 其中在南非和西班牙, 都被列为入侵性最强的等级。因此, 本研究通过这2个树种有性和无性繁殖体的数量、传播距离, 预测其入侵潜力, 以更好地保护当地的生态环境和生物资源。
1 调查地概况调查地点设在黑荆和银荆栽培较多的云南省楚雄彝族自治州东瓜镇和禄丰县和平乡。楚雄州位于25° 04′10″—25°13′42″N, 101°15′07″—102°12′04″E。属北亚热带季风气候区, 年均温15.5 ℃, 极端最高温33.4 ℃, 极端最低温-4.8 ℃。年平均降水量826.3 mm, 主要集中在5 —10月; 平均相对湿度72 %; 主导风向为西南风, 年平均风速1.7 m·s-1。土壤属于山地红壤, 砂壤土, 厚度20 ~ 100 cm, 颜色为黄红色至红色, 干燥, pH值4.6 ~ 6.5, 成土母质为紫砂页岩。调查林分与单株基本情况见表 1。
|
|

土壤种子库是指存在于土壤上层凋落物和土壤中全部存活种子的总和(Simpson, 1989)。调查于2004年3月初进行, 从水平和垂直2个方向调查黑荆和银荆林分及单株的土壤种子库。在林地内, 根据林分大小按样线法设4 m ×4 m的大样方, 样方间隔15 m以上, 每个大样方内划定面积为20 cm ×20 cm的小样方3个, 分枯落物层、0 ~ 3 cm层取样; 单株调查时在上风向、下风向、垂直风按树高0.5倍、1倍、2倍距离取样, 每个点取3个样。将土样带回, 用筛选法分出各层的黑荆或银荆的种子, 统计数量并计算标准误差, 林分内不同深度做方差显著性检验。
将土壤种子库中收集回的种子分为完整种子、空瘪粒种子、腐烂种子及动物损害种子, 然后用完整种子做发芽试验。发芽预处理按中国林木种子检验规程(GB2722 -1999)规定的方法进行, 即100 ℃水烫5 min, 然后冷水中浸泡24 h, 以冷水中浸泡24 h作对照, 滤纸为基质, 30 ℃恒温, 8 h光照, 4次重复, 每个重复100粒种子, 统计发芽率。
2.2 林缘与单株幼苗扩散林缘幼苗扩散调查在林缘开阔地按上风向、下风向和垂直风向置各设1 ~ 2个宽2 m的样方, 长度随幼苗的扩散距离而定, 到最远的幼苗为止。记录幼苗离林缘的距离、幼苗高度、株数, 刨根并分别实生苗和萌生苗统计。单株幼苗扩散调查选半径20 m内无相同树种的黑荆和银荆单株, 以母株为圆心, 沿上风向、下风向、垂直风向各取45°扇形调查, 如图 1所示, 记录幼苗离母株的距离、幼苗高度、株数, 也分实生苗和萌生苗统计。

|
图 1 单株扩散调查示意图 Fig. 1 Spread of an individual tree |
如表 2所示, 黑荆和银荆林分土壤种子库数量都很大, 达1 475 ~ 28 882粒·m-2; 黑荆2个林分的土壤种子库数量都大于银荆, 特别是东瓜镇黑荆林, 枯落物层和0 ~ 3 cm内数量都最多。种子在枯落物中分布较多, 达738 ~ 22 160粒·m-2, 在0 ~ 3 cm中分布较少, 只有738 ~ 6 722粒·m-2。方差分析表明:黑荆林分土壤种子库垂直分布差异显著(东瓜镇F =69.444 >F0.01 =8.096, 和平乡F =5.691 >F0.05 =4.001), 银荆林分不显著(F =0 < F0.05 =5.987), 说明土壤种子库数量大小除了与林分本身结实量不同有关外, 还与地表微环境有关。调查时发现, 黑荆林分内枯落物层较厚, 有利于种子散落后存留; 表层土壤深厚疏松, 有利于种子从枯落物中进入表层土壤中。银荆林分内坡度较大(30°以上), 地面植物呈团状分布, 种子散落后落入沟底随水流走, 不易在地表及土壤内存留, 所以土壤种子库数量较小, 而且分布不均。
|
|
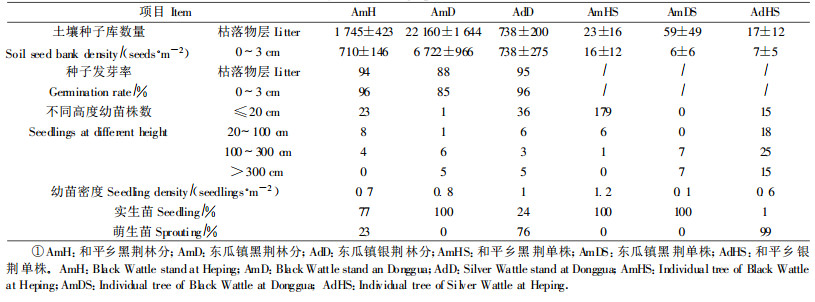
从表 2可以看出, 黑荆和银荆林分土壤种子库中的种子活力都很高, 经过沸水处理的种子发芽率分别为87 %、95 %、95 %, 冬瓜镇黑荆林分较低; 枯落物层与0 ~ 3 cm层内种子发芽率差别不大, 表明这2个树种种子长时间存在于土壤中生活力仍很高。未经沸水处理的黑荆种子发芽率很低, 只有6 %和1 %, 表明黑荆在自然条件下发芽很困难, 但也有自然萌发现象发生, 能够天然更新。
3.3 黑荆和银荆单株土壤种子库数量与空间结构调查结果表明, 林分密度也影响土壤种子库数量。表 2可以看出, 黑荆和银荆单株土壤种子库数量与其林分相比非常小, 仅有23 ~ 64粒·m-2, 枯落物层数量大于0 ~ 3 cm层, 但差别不明显。均值的标准误差较大, 说明各点间种子分布很不均匀, 离母株越远种子越少。土壤种子库的水平分布与风向关系密切, 风有助于种子传播。从图 2 a、b可知, 顺风向数量最大, 黑荆和银荆分别为350和650粒·m-2, 而垂直方向分别为200和25粒·m-2; 顺风向传播也较远, 是其他方向的2倍; 银荆单株土壤种子库最远可以分布到16 m, 黑荆最远20 m, 都约是树高的2倍, 为近距离传播。

|
图 2 黑荆与银荆单株土壤种子库 Fig. 2 Horizontal distribution of soil seed bank of individual trees in Black Wattle and Silver Wattle a.和平乡黑荆单株Individual tree of Black Wattle in Heping; b.和平乡银荆单株Individual tree of Silver Wattle in Heping. |
幼苗数量是树种繁殖对策与周围环境相互作用的结果。从表 2可以得出:银荆林缘幼苗扩散数量大于黑荆, 可达1.0株·m-2, 幼苗以根蘖苗为主, 占幼苗总数的76 %; 黑荆林缘幼苗扩散数量分别为0.7和0.8株·m-2, 以实生苗为主, 占分别总数的77 %和100 %。从高度分布来看, 和平乡黑荆林分和东瓜镇银荆林分苗高小于20 cm的小苗最多, 分别占幼苗总数的66 %和72 %, 东瓜镇黑荆幼苗在各个高度都有分布, 其中1 m以上大苗居多而小苗少。调查时发现, 苗高20 cm以下的幼苗多为当年生苗, 抵御环境干扰的能力非常弱, 1 m以上的大苗已经能够长成大树, 3 m以上的幼树基本都开花结实。从扩散距离来看, 黑荆和银荆林缘扩散相似, 最远距离都可达到25 m左右, 约是土壤种子库的分布的2倍。调查林分都有放牧干扰, 人或牲畜的活动可能使少量种子远距离传播而形成幼苗甚至大树。
3.4.2 单株幼苗扩散黑荆单株幼苗扩散同样也是以实生苗为主, 和平乡幼苗数量大于其林缘, 冬瓜镇反之, 林分郁闭度影响幼苗更新。从高度来看, 和平乡黑荆单株周围苗高20 cm以下幼苗占96 %, 与林缘扩散相比, 虽然幼苗密度大但真正能长成大树的比例却不高。东瓜镇黑荆单株周围幼苗只有0.1株·m-2, 但高度都在1 m以上。调查时发现, 冬瓜镇单株周围有蓝桉生长, 郁闭度可达0.8, 光照条件不利于幼苗的更新和生长。银荆单株也以根蘖为主, 周围幼苗数量有0.6株·m-2, 小于其林缘及黑荆单株, 但1 m以上的大苗数量多, 占周围幼苗总数的55 %。由此可以看出, 种子繁殖幼苗数量大, 但繁殖成功率不如根蘖繁殖高。
从图 3 a和b可以看出, 黑荆单株下风向幼苗数量最多, 和平乡和东瓜镇分别为1.9和0.2株·m-2, 其中和平乡单株3.5 m以内幼苗可达4.8株· m-2 (图 3 a); 此方向幼苗扩散距离也最远, 都在12 m以上, 也表明风力有助于黑荆种子的扩散。垂直风向幼苗数量也很多, 但因为是上坡, 扩散距离小。上风向数量较少, 和平乡和东瓜镇单株分别为0.2和0.1株·m-2, 因为是下坡, 扩散距离较远。可见种子也依靠重力传播, 地形对其影响也很大。从图 3 c可以看出, 银荆单株扩散没有明显的方向性, 随着距离的增加3个方向幼苗数量均增加, 且扩散数量相差不大, 上、下、垂直风向分别为0.6、0.8、0.5株·m-2, 最远距离为11.9 m, 较黑荆单株扩散稍近。

|
图 3 黑荆与银荆单株幼苗扩散 Fig. 3 Horizontal distribution of seedlings of individual trees of Black Wattle and Silver Wattle a.和平乡黑荆单株Individual tree of Black Wattle in Heping; b.冬瓜镇黑荆单株Individual tree of Black Wattle in Donggua; c.和平乡银荆单株Individual tree of Silver Wattle in Heping. |
生物入侵的前提就是有大量繁殖体的存在。银荆和黑荆都兼具有性和无性两种繁殖类型, 黑荆以种子繁殖为主, 繁殖体数量大, 银荆以根蘖繁殖为主, 繁殖成功率高。
土壤种子库是植物天然更新与植被恢复的物质基础(Moles et al., 1999), 可以揭示种群和群落动态。森林土壤种子库的密度一般在100 ~ 1 000粒·m-2(Halpern et al., 1999), 黑荆和银荆林分土壤种子库与其相比数量大得多, 黑荆林分最大可达28 882粒·m-2, 与热带森林土壤种子库相当(2.5万~ 3.0万粒·m-2)(曹敏等, 1999)。种子主要分布在枯落物层, 有利于种子继续传播; 小部分种子处于下层土壤中, 环境较为稳定, 种子可以存活较长的时间, 形成植物种群的天然基因库, 有助于种群的持续稳定(张咏梅等, 2003)。在南非, 柳叶相思(Acacia saligna)入侵灌丛植被, 长期入侵立地的土壤种子库数量达8 091粒·m-2, 新入侵立地有2 251粒·m-2, 降低了原有植物土壤种子库的数量, 严重影响当地的植被恢复(Holmes, 2002)。黑荆林分土壤种子库数量远大于它, 银荆林分与之接近, 因此具有较大的入侵潜力。
入侵性植物在竞争中通常倾向于采取“r -对策”, 即以非常高的生殖率和扩散能力取得生存机会(Reichard et al., 1997; Rejmanek et al., 1996), 黑荆和银荆土壤种子库数量大而幼苗数量相对较近, 正是这种生存对策。黑荆和银荆种子能够自然萌发, 但由于种皮致密坚硬, 表面有蜡质, 有机械休眠。这种特性使种子在土壤中存在多年仍有较高的活力, 保证种群顺利度过不良环境并在合适的条件下萌发成林。银荆也能产生大量种子, 但却以无性繁殖为主。无性繁殖能使种群快速增长, 土壤的破坏不会影响物种的续存, 相反还有助于其繁殖体的传播(Willis et al., 1999)。有研究表明, 银荆林冠下浅锄、割灌等干扰能促进林分天然更新(吴梅英, 2001)。种子的萌发和幼苗成活决定林下更新, 如果种子萌发并且成活, 遇到开阔林地, 种群就能迅速增长(Greenberg et al., 2001)。调查结果表明, 黑荆和银荆种群向外扩散速度慢, 但如果遇到合适的生境, 也能够迅速成林, 具有较强的入侵潜力。
4.2 扩散机制与扩散距离植物的长距离扩散决定入侵速度和分布范围(Neubert et al., 2000)。种子水平分布越广, 其扩散能力越强, 有利于种子迅速找到适宜的生存环境, 建立新的种群。金合欢属植物的豆荚富含纤维素, 果皮失水或吸水后表面产生不均匀的应力, 达到一定阀值时突然打开将种子弹射出去, 种子在重力的作用下落到地面, 再通过其他方式进行二次分布(New, 1984)。风力在黑荆和银荆种子传播过程中起重要作用, 下风向种子数量最大, 传播距离也最远, 黑荆幼苗扩散也是如此。Dalling等(1998)曾指出, 产生小粒种子的先锋树种, 种子传播距离一般都在30 m以上; Tackenberg等(2003)也总结出由风力传播的种子50 %超出20 m的范围, 20 %超出30 m。黑荆和银荆与其相比传播距离较近。种子被风远距离传播决定于天气和自身下落速度(Horn et al., 2001), 而自身下落速度与种子结构和树冠高度有关。黑荆和银荆树高约10 ~ 15 m, 种子小, 不具备风力传播的特点, 如种子具翅、气囊等, 被风远距离传播的可能性不大。在澳大利亚和非洲, 黑荆和银荆的种子可以被动物传播, 如蚂蚁和一些食草动物(New, 1984)。在我国, 据观察, 栽培地区牛羊等牲畜吃其枝叶豆荚的情况不多, 被大型动物传播的几率很小。种子被蚂蚁等小动物移动的可能性存在, 调查时也发现小堆饱满的种子聚集在一起, 但也只是近距离传播。银荆通过根蘖繁殖, 幼苗扩散距离取决于根系的分布, 较种子传播距离近。
4.3 入侵防范与利用黑荆和银荆土壤种子库数量大, 根部萌蘖能力强, 适应性广, 是荒山绿化和水土保持的好树种, 是当地重要的造林树种。但其具有较强的入侵潜力, 在引种栽培时要因地适宜, 加强管理。在我国, 黑荆和银荆都是近距离的扩散。但在南非, 其种子随流水传播, 已经造成巨大危害(Maitre et al., 2002)。因此, 将黑荆和银荆种植于沟渠旁用于水土保持时要慎重, 谨防其繁殖成灾, 破坏原有的生态环境。黑荆和银荆在不同地区之间的扩散主要是人为引种栽培造成的, 在我国远距离自然扩散的现象还未见报道, 因此, 避免在当地特有群落或珍稀濒危物种附近栽种, 以免发生入侵而造成无法挽回的后果。黑荆和银荆有很强的抗干扰能力, 火烧能够促进种子萌发及根部萌蘖, 机械防除不能有效控制。但种子萌发和幼苗生长都需要开阔地和充足的光照, 因此, 栽植时较高的林分密度有利于限制其林下的天然更新, 从而降低种群增长。
曹敏, 唐勇, 张建侯, 等. 1999. 西双版纳热带雨林土壤种子库与地面植被的关系. 应用生态学报, 10(3): 279-282. DOI:10.3321/j.issn:1001-9332.1999.03.006 |
潘志刚, 游应天. 1994. 中国主要外来树种引种栽培. 北京: 科学技术出版社.
|
吴梅英. 2001. 银荆林冠下天然更新的研究. 福建林业科技, 28(2): 32-34. DOI:10.3969/j.issn.1002-7351.2001.02.009 |
徐汝梅. 2003. 生物入侵———数据集成、数量分析与预警. 北京: 科学出版社.
|
徐汝梅, 叶万辉. 2003. 生物入侵———理论与实践. 北京: 科学出版社.
|
徐承远, 张文驹, 卢宝荣, 等. 2001. 生物入侵机制的研究. 生物多样性, 9(4): 430-438. DOI:10.3321/j.issn:1005-0094.2001.04.016 |
张咏梅, 何静, 潘开文, 等. 2003. 土壤种子库对原有植被恢复的贡献. 应用与环境生物学报, 9(3): 326-332. DOI:10.3321/j.issn:1006-687X.2003.03.025 |
国家质量监督局.2000.GB2722-1999, 林木种子检验规程.北京: 中国标准出版社
|
Barret S C H. 1983. Crop mimicry in weeds. Economic of Botany, 37: 255-282. DOI:10.1007/BF02858881 |
Chapin F S, Zavaleta E S, Eviner V T, et al. 2000. Consequences of changing biodiversity. Nature, 405: 234-242. DOI:10.1038/35012241 |
Dalling J W, Swaine M D, Garwood N C. 1998. Dispersal patterns and seed bank dynamics of pioneer trees in moist tropical forest. Ecology, 79: 564-578. DOI:10.1890/0012-9658(1998)079[0564:DPASBD]2.0.CO;2 |
Greenberg C H, Smith L M, Levey D J. 2001. Fruit fate, seed germination and growth of an invasive vine:an experimental test of sit and wait strategy. Biological invasions, 3: 363-372. DOI:10.1023/A:1015857721486 |
Halpern C B, Evans S A, Nielson S. 1999. Soil seed banks in young, close-canopy forests of the Olympic Peninsyla, Washington:potential contributions to understory reinitiation. Canadian Journal of Botany, 77: 922-955. |
Hendersong L, Musil K J. 1984. Exotic woody plant invaders of the Transvaal. Bothalia, 15: 297-313. |
Holmes P M. 2002. Depth distribution and composition of seed-banks in alien-invaded and uninvaded fynbos vegetation. Austral Ecology, 27: 110-120. DOI:10.1046/j.1442-9993.2002.01164.x |
Horn H S, Nathan R, Kaplan S R. 2001. Long-distance dispersal of tree seeds by wind. Ecological Research, 16: 877-885. DOI:10.1046/j.1440-1703.2001.00456.x |
Lowe S, Browne M, Boudjelas S et al.2000.100 of the World' s Worst Invasive Alien Species. www.Issg.org booklet.pdf
|
Maitre D C L, Wilgen B W V, Gelderblom C M, et al. 2002. Invasive alien trees and water resources in South Africa :case studies of the costs and benefits of management. Forest Ecology and Management, 160: 143-159. DOI:10.1016/S0378-1127(01)00474-1 |
Moles A T, Drake D R. 1999. Potential contribution of the seed rain and seed bank to regeneration of native forest under plantation pine in New Zealand. New Zealand Journal of Botany, 37: 83-93. DOI:10.1080/0028825X.1999.9512615 |
Neubert M G, Caswell H. 2000. Demography and dispersal:Calculation and sensitivity analysis of invasion speed for structured populations. Ecology, 81: 1613-1628. DOI:10.1890/0012-9658(2000)081[1613:DADCAS]2.0.CO;2 |
New T R. 1984. Biology of Acacias. Oxford University Press.
|
Nyoka B I.2003.Biosecurity in Forestry : A Case Study on the Status of Invasive Forest Tree Species in Southern Africa.Forest Biosecurity Working Papers, FAO, Roman
|
Poschlod P, Kleyer M, Tackenberg O. 2000. Databases on life history traits as a tool for risk assessment in plant species. Zeitschrift fur Oekologie und Naturschutz, 9: 3-18. |
Quentin C B, Fuller J L. 1995. Plant invaders:the threat to natural ecosystem. Chapman & Hall.
|
Reichard S H, Hamilton C W. 1997. Predicting invasions of woody plant introduced into North America. Conservation Biology, 11: 193-203. DOI:10.1046/j.1523-1739.1997.95473.x |
Rejmanek M, Richardson D. 1996. What attributes make some plant species more invasive?. Ecology, 77(6): 1655-1661. DOI:10.2307/2265768 |
Simpson R L. 1989. Ecology of soil seed bank. San Diego :Academic Press: 149-209. |
Smith C W.1985.Impact of alien plants on Hawaii' s native biota, in Hawaii' s Terrestrial Ecosystems: preservation and management.In: Stone C P and Scott J M.Cooperative National Park Resources Studies Unit, University of Hawaii, Honolulu
|
Tackenberg O, Poschlod P, Bonn S. 2003. Assessment of wind dispersal of potential in plant species. Ecological Monographs, 73(2): 191-205. DOI:10.1890/0012-9615(2003)073[0191:AOWDPI]2.0.CO;2 |
Vitousek P M, D'Antonio C M, Loope L L, et al. 1997. Introduced species:a significant component of human-caused global change. New Zealand Journal of Ecology, 21: 1-16. |
Wilcove D S, Rothstein D, Dubow J, et al. 1998. Quantifying threats to imperiled species in the United States. Bioscience, 48: 607-615. DOI:10.2307/1313420 |
Willis A J, Thomas M B, Lawton J H. 1999. Is the increased vigor of invasive weeds explained by a trade-off between growth and herbivore resistance?. Oecologia, 120: 632-640. DOI:10.1007/s004420050899 |