 2006, Vol. 42
2006, Vol. 42文章信息
- 何恒斌, 张惠娟, 贾桂霞.
- He Hengbin, Zhang Huijuan, Jia Guixia.
- 磴口县沙冬青种群结构和空间分布格局的研究
- Population Structure and Spatial Distribution Pattern of Ammopiptanthus mongolicus in Dengkou County, Inner Mongolia Autonomous Region
- 林业科学, 2006, 42(10): 13-18.
- Scientia Silvae Sinicae, 2006, 42(10): 13-18.
-
文章历史
- 收稿日期:2005-09-27
-
作者相关文章



2. 内蒙古巴彦淖尔盟林业局 临河 015000
2. Bayannaoer League Forestry Bureau, Inner Mongolia Linhe 015000
沙冬青(Ammopiptanthus mongolicus)是生长在西北荒漠地区的常绿阔叶灌木, 第三纪孑遗种, 为国家首批重点保护的珍稀濒危植物(刘果厚, 1998)。沙冬青是优良的固沙植物, 具有抗热、抗旱、抗寒、耐盐碱、耐贫瘠、耐沙埋、抗风蚀等特性(中国植物志编辑委员会, 1997), 且地上部分分枝多、冠幅较大、阻沙能力强。不少学者对沙冬青的生物学特征、濒危原因、抗逆生理、生理生态、超微结构、生物化学、种群等方面做了大量的研究(孙祥等, 1994; 王庆锁等, 1995; 周宜君等, 2001; 陈曦等, 2002; 费云标等, 1994; 尉秋实等, 2005)。对内蒙古磴口县不同生境沙冬青种群在不同尺度上的空间分布格局进行分析, 旨在阐明种群空间分布格局与生境、种群年龄结构之间的动态关系, 揭示种群发育的空间动态与生物学机制, 为阐明沙冬青种群的繁殖扩散机制、种群更新及群落演替规律提供理论依据, 为制定科学的保护措施和合理的开发利用提供依据。
1 研究区概况磴口县隶属于内蒙古巴彦淖尔盟, 处于著名的河套平原产粮灌溉区内(40°13′ N, 107°05′ E)。气候较温暖湿润, 年均降水量149.2 mm, 年均蒸发量2 380.6 mm, 年均相对湿度46%, 地下水位较高。土壤区系属漠钙土带(王葆芳等, 1998), 土壤主要类型为粉沙粘土和风沙土。属荒漠与干草原的过渡地带, 植被类型多样。磴口县天然分布有沙冬青, 是沙冬青的重要分布区之一。
2 研究方法 2.1 取样方法与野外调查2005年6月和7月, 综合磴口县土壤类型、水分等条件, 选取3个沙冬青群落典型样地, 样地概况见表 1。在样地1设置1个50 m×50 m的样地, 在样地2设置1个20 m×100 m的样地, 在样地3设置1个30 m×50 m的样地(受地形限制, 3个样地面积有差异); 每个样地划分为5 m×5 m的小样方, 共设小样方240个。调查记录每个样方内沙冬青的树高(H)和冠幅(W), 以及其他物种的种名、株高、盖度、株数和物候期等; 并记录每个样地的生境因子如海拔、坡向、坡度、坡位、土壤类型等。以样地的一条边为X轴, 以其偏东向的垂直边作为Y轴, 记录每一株沙冬青的坐标值。工作于2005年8月完成。
|
|

在侯平等(1994)和尉秋实等(2005)研究结果的基础上, 结合沙冬青种群在自然环境下的生长特点, 对树高和冠幅进行分级。以20 cm为一个树高级(从0开始共分为10级, 由小到大的顺序为a、b、c、d、e、f、g、h、i和j, 见图 1), 30 cm为一个冠幅级(从0开始共分为14级, 由小到大的顺序为a、b、c、d、e、f、g、h、I、j、k、l、m和n, 见图 2), 统计样地内各高度级和幅度级的个体数量。年龄结构是反映种群动态的一个重要指标(谢宗强等, 1999), 为了避免破坏珍稀濒危植物, 目前多采用单株的大小结构(乔木树种多采用径级)代替年龄结构(闫桂琴等, 2001; 谢宗强等, 1995)。沙冬青属于灌木, 基部分枝多, 无明显主干, 年轮不易辨别, 应用生长锥或径级研究种群动态的难度较大(尉秋实等, 2005)。结合前人的研究成果和沙冬青种群在自然条件下的生长特性, 应用天然沙冬青种群的个体树高和冠幅分析种群动态。

|
图 1 各树高级沙冬青密度 Fig. 1 Densities of different tree height grades |

|
图 2 各冠幅级沙冬青密度 Fig. 2 Densities of different width grades |
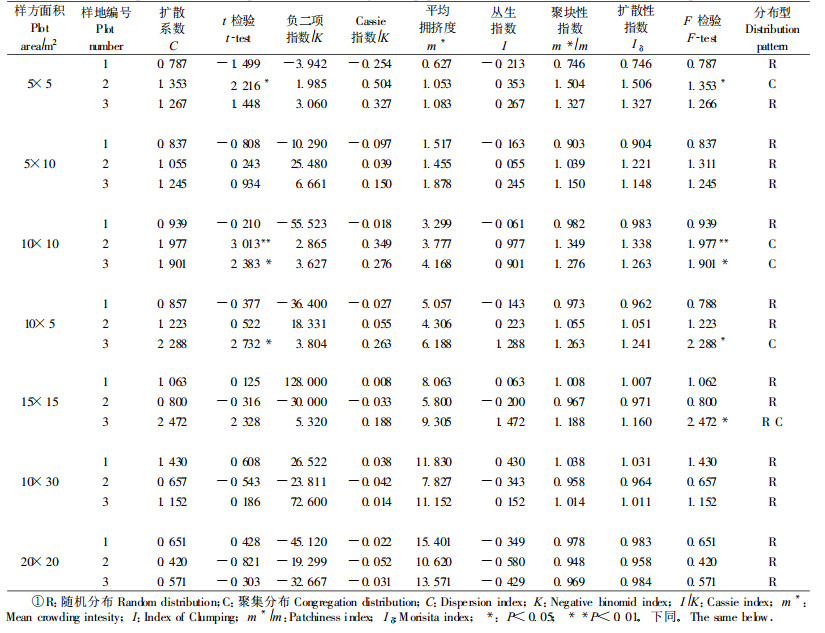
以样地内沙冬青个体定位数据为依据, 分5 m×5 m、5 m×10 m、10 m×10 m等7种样方格子分别进行统计分析; 应用扩散系数C (C值用t检验, )、负二项指数(K)、平均拥挤度(m*)、聚块性指数(m*/m)、扩散型指数Iδ (Iδ值用F检验), 丛生指数(I)和Cassie指数(1/K)判定沙冬青种群的格局分布类型并测定其聚集强度, 分析种群不同年龄级的空间分布格局(李海涛, 1995; 郑元润, 1997; 张金屯, 2004; Greig-Smith, 1983)。
3 结果与分析 3.1 沙冬青种群结构由图 1、2可以看出, 3个样地均表现为缺乏幼苗(H≤20 cm, W≤30 cm)和幼树(20 cm < H≤40 cm, 30 cm < W≤60 cm), 中龄(60 cm < H≤120 cm, 60 cm < W≤180 cm)和老龄植株(H > 120 cm, W > 180 cm)较多。样地1种群分布极不均匀, 没有幼苗, 幼树极少, 中龄植株所占比例最大, 老龄植株较少, 说明在过去一段时间内种群呈现了增长趋势, 但由于近年来生境的改变, 种群的自然更新存在很大的困难, 目前已经出现了断代现象, 老龄植株也濒于死亡。样地2种群自然更新状况相对较好, 存在一定量的幼苗和幼树, 由植株的树高和冠幅反映出来的生长状况也较好, 说明样地2相对较适合沙冬青的更新和发育, 但年龄结构仍呈现中间大两头小的格局。样地3种群也呈现幼龄植株少, 中龄和老龄植株较多的趋势, 而且老龄植株所占比例有所增多。
3.2 种群空间分布格局 3.2.1 不同生境下沙冬青种群的空间分布格局类型和格局强度环境的空间异质性, 是影响种群空间分布格局形成和发展的主要因素之一, 随着时间的变化, 植物的分布格局也发生改变。对不同生境沙冬青种群空间分布类型和聚集强度进行分析(表 2), 结果表明:各项聚集度指标值一致, 样地2在25和100 m2集群分布, 对C值进行t检验分别达到显著(2.216)和极显著水平(3.013), 对Iδ进行F检验也分别达到显著(1.353)和极显著水平(1.977)。样地3在150 m2呈集群分布, 进行t和F检验达到显著水平; 而在225 m2范围内, 对C值进行t检验显示呈随机分布, 对Iδ进行F检验表明种群呈聚集分布。样地1在7个取样面积下, 均呈随机分布。将样地中的种群个体进行定位, 按7种不同大小样方进行取样, 分析种群在不同取样尺度下的格局强度(表 2)。沙冬青种群的格局强度随取样尺度的变化而变化, 且不同生境沙冬青种群的格局强度随尺度变化而表现出来的变化趋势基本一致, 呈现“小→大→小”的趋势。但格局强度最大值和出现最大值时的面积不同, 样地1在300 m2有最大聚集强度(C=1.430, Iδ=1.031), 但呈随机分布; 样地2在100 m2有最大聚集强度(C=1.977, Iδ=1.338), 呈聚集分布; 样地3在150 m2有最大聚集强度(C=2.288, Iδ=0.241), 呈聚集分布。
|
|
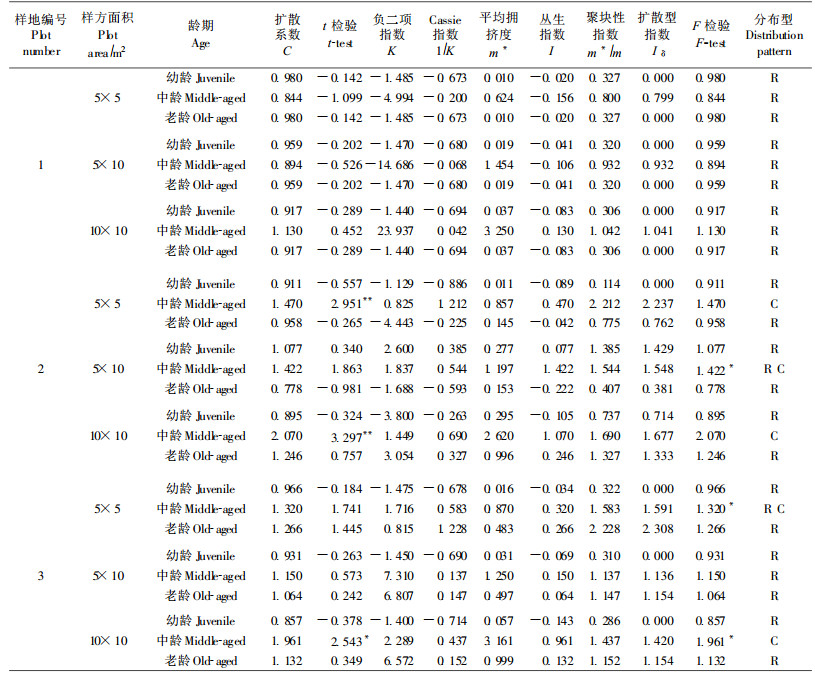
种群的空间分布格局是指种群个体某一时刻在空间的散布状态。它既有时间上的限制, 又有可能随时间序列而演变(宋萍等, 2005)。分析种群在不同发育阶段的空间格局变化可以推断种群新生个体的产生、成年个体的死亡以及种群遭受人为干扰的概况(蔡飞等, 1997; 张文辉, 1998)。沙冬青种群幼龄、中龄和老龄植株的空间分布类型因生境不同而有所差异(表 3)。各样地内不同发育阶段种群的分布格局大体上表现为:幼龄和老龄植株均成随机分布, 中龄植株呈聚集分布。
|
|
比较不同种群树高和冠幅结构, 能较好体现环境因子对种群的作用强度和种群的适应特征。不同沙冬青种群在树高和冠幅上差异显著, 生长于黄河故道的沙冬青种群平均树高和冠幅均高于其他2个生境下的种群, 说明该生境条件和群落结构更适合沙冬青的生长。这与其他植物在其适生的环境条件下能够充分地利用光、温、水、土壤因素, 实现自身生存扩展最大化的特征基本一致(陈劲松等, 2001; 殷淑燕等, 1997; 张文辉等, 2002)。
不同沙冬青种群环境和立地条件不同, 生长发育明显不同。对不同种群的年龄结构进行分析显示, 虽然种群间和种群内最大年龄有差异, 但年龄级的分布趋势却相近。一般濒危植物种群的年龄结构多呈衰退型, 但在适合的生境条件下呈进展型, 并且在自然条件下能够实现自我更新(张文辉等, 2002), 但磴口县3个沙冬青种群均表现出幼龄植株少、中老龄植株多的结构, 说明磴口县沙冬青种群扩展能力弱化的形势相当严重, 均出现了严重的断代现象, 这与尉秋实等(2005)关于阿拉善左旗沙冬青种群的报道一致。沙冬青种群分布面积只局限于内蒙古、宁夏等的少数地区。由于环境的自然间断和人为破碎化, 沙冬青在其分布地形成一定规模的聚集斑块, 这种分布格局特征可能是对环境异质性的响应和种内竞争的结果。在某种意义上, 种群的空间分布格局是种群对环境长期适应的结果, 因而种群格局通常反映着一定环境因子对个体行为、生存和生长的影响(马克明等, 2000)。因此, 研究样地中各种环境因素, 如地形、土壤厚度、土壤养分、土壤含水量和地下岩石组成等对沙冬青种群格局的具体影响及对应关系, 有助于进一步解释种群个体分布格局的成因(李海涛, 1995)。
一般情况下, 植物以母株为中心的繁殖方式使低龄树呈现聚集分布; 随着年龄的增加, 树木对光因子和营养条件的要求加大及自疏、它疏等作用, 种群会趋于均匀分布; 随着年龄的继续增加, 种群将趋于随机分布(郭华等, 2005)。而沙冬青种群的空间分布格局却呈现出不同于一般的特点:幼龄和老龄植株呈随机分布, 中龄植株呈聚集分布。这种格局动态可能是沙冬青特殊的生物学特性和环境因子共同作用的结果。研究表明, 由于环境破坏, 生物多样性减少, 原有群落中生物间的互惠互利的稳定关系已经破坏, 对植物产生有利影响的昆虫、鸟类和其他动物、微生物数量减少, 而对濒危植物产生不利影响的动物和微生物活动增强(张文辉等, 2002)。根据刘果厚(1998)的报道, 虽然沙冬青结实量较大, 但种子被虫蛀食率较高, 抽测20个样方的种子, 平均虫蛀率为30%, 因而减少了天然更新的繁殖材料。在实验室条件下, 沙冬青的发芽率可达到70%, 但幼苗的存活率较低(丁琼, 2004)。沙冬青分布区严酷的自然条件, 又进一步限制了本已较少的沙冬青种子的萌发和幼苗的存活。多个环节的叠加作用使沙冬青种群的幼龄植株极少甚至没有, 从而使沙冬青幼龄植株只零星地分布于种群中, 表现为随机分布。随着年龄的增长, 沙冬青植株的抗逆性加强, 对环境的适应能力增强, 而且不同世代的幼苗逐渐成为中龄植株, 使中龄植株在适合沙冬青生长发育的局部环境中数量有所增加, 表现出一定程度的聚集分布。进入老龄期, 不仅有种间和种内的竞争, 而且生理逐渐衰老, 在环境筛的作用下个体死亡, 种群聚集度下降, 表现为随机分布。因此, 沙冬青分布区严酷的环境条件主导了沙冬青种群格局的形成, 并进一步影响了种群格局的动态变化。
蔡飞, 宋永昌. 1997. 武夷山木荷种群机构和动态的研究. 植物生态学报, 21(2): 138-148. DOI:10.3321/j.issn:1005-264X.1997.02.005 |
陈劲松, 苏智先. 2001. 缙云山马尾松种群生物量生殖配置研究. 植物生态学报, 25(6): 704-708. DOI:10.3321/j.issn:1005-264X.2001.06.010 |
陈曦, 卢存福, 蒋湘宁, 等. 2002. 植物抗冻蛋白及其基因工程研究的新进展. 北京林业大学学报, 24(3): 94-98. DOI:10.3321/j.issn:1000-1522.2002.03.020 |
丁琼. 2004. 共生菌在濒危植物沙冬青引种栽培中应用研究. 北京: 北京林业大学, 33-41.
|
费云标, 孙龙华, 黄涛, 等. 1994. 沙冬青高活性抗冻蛋白的分离与鉴别. 植物学报, 36(8): 649-650. |
郭华, 王孝安, 肖娅萍. 2005. 秦岭太白红杉种群空间分布格局动态及分形特征研究. 应用生态学报, 16(2): 227-232. DOI:10.3321/j.issn:1001-9332.2005.02.007 |
侯平, 尹林克, 严成. 1994. 沙冬青生物量的研究. 干旱区研究, 11(1): 16-22. |
李海涛. 1995. 植物种群分布格局研究概况. 植物学通报, 12(2): 19-26. |
刘果厚. 1998. 阿拉善荒漠特有植物沙冬青濒危原因的研究. 植物研究, 18(3): 341-345. |
马克明, 祖元刚. 2000. 兴安落叶松种群格局的分形特征记盒维数. 植物研究, 20(1): 104-111. DOI:10.3969/j.issn.1673-5102.2000.01.018 |
宋萍, 洪伟, 吴承祯, 等. 2005. 珍稀濒危植物桫椤种群结构与动态研究. 应用生态学报, 16(3): 413-418. DOI:10.3321/j.issn:1001-9332.2005.03.004 |
孙祥, 于卓. 1994. 沙冬青根系的研究. 干旱区研究, 11(1): 53-56. |
王葆芳, 熊士平, 任培政. 1998. 乌兰布和沙地新开发人工绿洲防护林体系综合效益评价. 林业科学, 34(6): 12-21. |
王庆锁, 李勇, 张灵芝. 1995. 珍稀濒危植物沙冬青研究概况. 生物多样性, 3(3): 153-156. DOI:10.3321/j.issn:1005-0094.1995.03.007 |
谢宗强, 陈伟烈, 江明喜, 等. 1995. 八面山银杉林种群的初步研究. 植物学报, 37(1): 58-65. |
谢宗强, 陈伟烈, 路鹏, 等. 1999. 濒危植物银杉的种群统计与年龄结构. 生态学报, 19(4): 523-528. DOI:10.3321/j.issn:1000-0933.1999.04.017 |
闫桂琴, 赵桂仿, 胡正海, 等. 2001. 秦岭太白红杉种群结构与动态的研究. 应用生态学报, 12(6): 824-828. DOI:10.3321/j.issn:1001-9332.2001.06.006 |
殷淑燕, 刘玉成. 1997. 大头茶构件种群生物量及叶面积动态. 植物生态学报, 21(1): 83-89. DOI:10.3321/j.issn:1005-264X.1997.01.012 |
尉秋实, 王继和, 李昌龙, 等. 2005. 不同生境条件下沙冬青种群分布格局与特征的初步研究. 植物生态学报, 29(4): 591-598. DOI:10.3321/j.issn:1005-264X.2005.04.010 |
张金屯. 2004. 数量生态学. 北京: 科学出版社, 243-266.
|
张文辉. 1998. 裂叶沙参种群生态学. 哈尔滨: 东北林业大学出版社, 56-157.
|
张文辉, 祖元刚, 刘国彬. 2002. 十种濒危植物的种群生态学特征及致危因素分析. 生态学报, 22(9): 1512-1520. DOI:10.3321/j.issn:1000-0933.2002.09.021 |
郑元润. 1997. 不同方法在沙地云杉种群分布格局分析中的适用性研究. 植物生态学报, 21(5): 480-484. DOI:10.3321/j.issn:1005-264X.1997.05.010 |
中国植物志编辑委员会. 1997. 中国植物志. 北京: 科学出版社.
|
周宜君, 刘春兰, 冯金朝. 2001. 沙冬青抗旱、抗寒机理的研究进展. 中国沙漠, 21(3): 312-316. DOI:10.3321/j.issn:1000-694X.2001.03.020 |
Greig-Smith P. 1983. Quantitative plant ecology. Oxford: Blackwell, 68-97.
|