 2006, Vol. 42
2006, Vol. 42文章信息
- 罗明武, 邓柳红.
- Luo Mingwu, Deng Liuhong.
- 巴西橡胶树产胶与排胶机制研究进展
- Advances in Mechanisms of Latex Regeneration and Latex Flow of Hevea brasiliensis
- 林业科学, 2006, 42(9): 127-130.
- Scientia Silvae Sinicae, 2006, 42(9): 127-130.
-
文章历史
- 收稿日期:2005-01-31
-
作者相关文章



2. 中国热带农业科学院热带生物技术研究所热带作物国家重点实验室 海口 571101
2. State Key Laboratory of Tropical Crop Biotechnology the Institute of Bioscience and Biotechnology Chinese Academy of Tropical Agricultural Sciences Haikou 571101
天然橡胶是产胶植物的一种次生代谢物, 具有耐高温、高弹性等突出优点, 是合成橡胶所不可比拟的一种世界性工业原料和重要的战略物质, 具有极高的经济价值及应用前景。目前中国自产的天然橡胶远远不能满足消费的需求, 有50%以上需要进口来弥补。天然橡胶主要来源于大戟科的巴西橡胶树(Hevea brasiliensis), 该植物因经济寿命长、易栽培采割、胶质好、产胶量多而在热带及亚热带地区广泛种植。随着对天然橡胶需求的日益增长, 如何进一步提高橡胶树的产量已成为天然橡胶行业亟待解决的重大课题, 在传统的技术与方法基础上, 急待探索新的途径。
巴西橡胶树靠分布于树体内的乳管系统产胶, 当割胶切断乳管时, 受到乳管内液体膨压的作用, 乳管内的胞质除细胞核、线粒体外开始流出, 同时乳管周围薄壁细胞的水分渗入, 使胶乳稀释, 维持继续排胶。流出物含有较多的橡胶粒子, 呈淡黄至乳白色的乳状液体, 称为胶乳。随排胶时间延长, 排胶速度变慢, 一般经过一段时间排胶后, 由于特定的机制使割口胶乳凝固堵塞, 乳管细胞靠留下的细胞核、线粒体等重新合成新的内含物。2次割胶之间的胶乳再生能力及排胶时间是胶乳产量的主要限制因子(何康等, 1987)。因此, 研究巴西橡胶树产胶与排胶机制对于如何提高天然橡胶的产量及橡胶树转基因高产育种具有重要的意义。
1 巴西橡胶树产胶机制的研究进展巴西橡胶树的乳管系统耗费大量的能量合成天然橡胶, 这一合成过程是怎样进行和调控的呢?目前人们正试图从形态结构、生理及分子生物学等方面研究来揭开这一秘密。
1.1 巴西橡胶树产胶形态结构及生理学研究橡胶树产胶的功能靠分布于树体内的乳管系统来进行, 同龄树乳管列数的多少是橡胶树品系的特征, 和产胶量成正相关。割胶、排胶及茉莉酸均能促进乳管分化(郝秉中等, 1982;1984; Hao et al., 2000; Wu et al., 2002)。从乳管中排出的胶乳是乳管细胞原生质体的一部分, 它除了含有一般细胞的许多结构成分外, 还包括了3种特殊的结构:橡胶粒子、黄色体小泡和FW复合体(Frey-Wyssling粒子)。橡胶粒子占胶乳质量的20%~40%, 是由异戊二烯单体顺式聚合而成的长链烃。黄色体小泡类似于植物的液胞和动物细胞中的溶酶体, 内有胞液, 称为B乳清。黄色体中含有丰富的蛋白质、酶和Ca、Mg等二价阳离子, 这些蛋白质、酶类以及二价阳离子调节黄色体内外电势差, 维持着黄色体和胶乳的稳定(Cardosa et al., 1994; D'Auzac et al., 1995)。FW复合体也是乳管特有的、类似于质体的细胞器。
目前橡胶的生物合成途径已大致了解:从蔗糖开始, 经过糖酵解, 三羧酸循环生成乙酰辅酶A。利用ATP为能量, 以磷酸戊糖途径产生的NADPH为还原剂, 由乙酰辅酶A转变为甲羟戊酸, 再由甲羟戊酸转变成异戊烯焦磷酸(IPP), IPP在IPP异构酶催化下生成二甲基丙烯基焦磷酸(DMAPP)。然后IPP在反式异戊二烯转移酶(法尼二磷酸合成酶)催化下, 将IPPs加在DMAPP上生成C10、C15等异戊二烯类化合物。最后, 在橡胶转移酶催化下, IPP添加到异戊二烯类化合物上形成橡胶。在橡胶生物合成过程中, 橡胶分子的从头合成除需要IPP外, 还需要1分子烯丙基焦磷酸(如DMAPP、GPP、FPP、GGPP等)作为起始物(20-22), 它们同样也是植物体内其他类异戊二烯化合物如植物激素(GA、ABA)、甾体及萜烯类合成的前体。所不同的是橡胶生物合成的最后一步, 它由产胶植物独有的酶-橡胶转移酶催化。因此橡胶转移酶的鉴定和调控的研究是阐明植物体内橡胶生物合成调控机制的关键所在(D'Auzac et al., 1997)。
1.2 巴西橡胶树产胶分子生物学研究Kush等(1990)研究发现与橡胶合成相关的基因及与植物防卫相关基因在胶乳中高表达, 乳管存在特异表达基因。为了找出与橡胶生物合成相关的基因(如橡胶合成关键酶基因), Han及Ko等研究小组分别通过差减杂交、EST和cDNA-AFLP, 构建了胶乳差减文库, 获得了一些差异片段(Han et al., 2000; Jae-Heung et al., 2003)。目前已克隆的与产胶相关的基因在表 1列出。
|
|
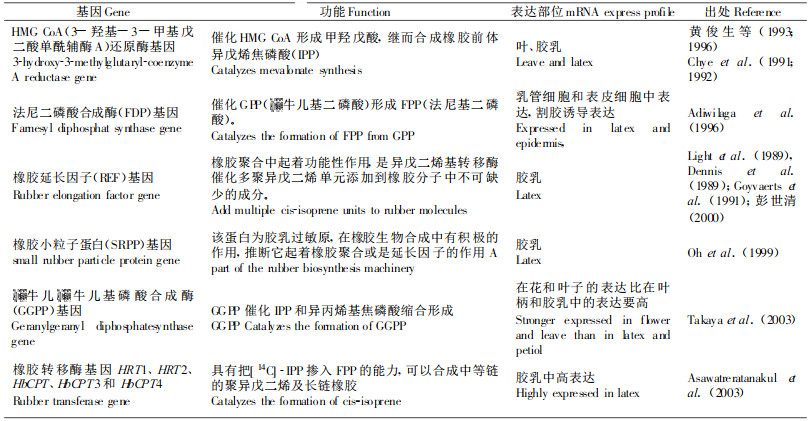
目前有关橡胶的生物合成途径已比较清楚, 也克隆了一些与产胶相关的功能基因, 但是合成过程是怎样调控的, 特别是有关橡胶生物合成的关键酶——橡胶转移酶的调控研究目前仍很少报道。另外与产胶相关的功能基因之间是怎样相互协调作用的, 乳管分化的分子机制是怎样的等问题仍不甚清楚。
2 巴西橡胶树排胶机制的研究排胶对胶乳产量的影响主要表现在胶乳的流速和延续时间2方面, 特别是胶乳流的延续时间。胶乳的停排是由于乳管割口处胶乳絮凝堵塞而引起的, 就此还提出了一些解释排胶的机理假说和体现排胶的生理指标(如排胶初速度, 堵塞指数, 硫醇R-SH含量、破裂指数等)。胶乳凝固假说主要有酶假说、静电中和假说和凝集假说。目前倾向于Gidrol等(1994)提出的“凝集素假说”, 该假说与几丁质酶的研究密切相关。即割胶后乳管伤口处或死皮发生的原位乳管内, 作为凝固因子的Hevein同Ca2+从黄色体释放到乳清中, Hevein和橡胶粒子膜上一种22 Kda蛋白(Hevein受体)通过N-乙酰葡糖胺结合, 形成多价交联而使胶乳凝固, 而在正常胶乳稳定情况下, 作为交联因子的Hevein大部分被隔离在黄色体中, 而作为22 KDa糖蛋白的糖基可以被几丁质酶切下, 游离糖基又能使橡胶粒子结合位点饱和, 结果导致橡胶素不能与橡胶粒子结合, 因而胶乳的凝固被抑制。
3 割胶及乙烯刺激对产胶排胶的影响割胶及乙烯利刺激均可以在一定程度上提高胶乳产量。特别是乙烯利作为化学刺激剂应用于提高胶乳产量以来, 增产效果非常明显, 但它们增加产量的机制至今仍不清楚。目前研究认为乙烯利对橡胶树的作用, 是促进排胶, 影响胶乳的代谢(Amalon et al., 1992; Gidrol et al., 1994)诱导胶乳细胞里一些特异基因的表达(Broekaert et al., 1990; Miao et al., 1993; Pujade-Renaud et al, 1994; Sivasubramaniam et al., 1995);外源乙烯不能诱导与橡胶生物合成相关的基因表达, 而割胶却可以诱导与橡胶生物合成相关的基因表达(Adiwilaga et al., 1996)。推测可能在未割树中, 胶乳细胞处在一种静止状态, 只是一些正常的持家基因转录物, 而割胶损失了大量的胶乳细胞质, 因此需要在乳管内建立一种新的代谢平衡, 来补充每次割胶后损失的细胞质(包括蛋白), 这时与橡胶生物合成相关的基因会过量表达。另外, 割胶诱导受伤树皮组织释放“伤乙烯”或外源乙烯刺激, 也会启动乳管内一些基因的过量表达, 影响胶乳的代谢, 促进胶乳排胶, 从而增加产量。但是, 过度割胶及乙烯刺激易使产胶排胶生理平衡失调, 导致橡胶树死皮病(TPD)发生, 进而乳管死亡无胶。
橡胶树的产胶排胶是一个动态的平衡系统, 是一个特殊而复杂的生理过程。产胶与排胶之间互相协调制约, 胶乳再生能力强才能为排胶提供基础, 而排胶顺畅、排胶量大又促使胶乳再生。另外产胶排胶还受到树体长势的影响, 如乳管蔗糖储备丰富, 蔗糖转化酶活性高, 橡胶合成原料充足等。总之, 如何协调好橡胶树的产胶排胶对于巴西橡胶树的种植, 橡胶产量的提高具有极其重大的意义。
郝秉中, 吴继林. 1982. 创伤(割胶)对橡胶乳管分化的影响. 植物学报, 24: 388-391. |
郝秉中, 吴继林, 云翠英. 1984. 排胶对橡胶树乳管分化的促进作用. 热带作物学报, 5(2): 19-23. |
何康, 黄宗道. 1987. 热带北缘橡胶树栽培. 广州: 广东科技出版社.
|
黄俊生, 孔德謇, 符少萍. 1993. 巴西橡胶HMG-CoA还原酶基因cDNA的扩增与克隆. 热带作物学报, 14(2): 11-15. |
黄俊生, 孔德謇. 1996. 橡胶树HMG-CoA还原酶基因结构序列分析. 热带作物学报, 17(2): 5-10. |
彭世清, 吴坤鑫, 陈守才. 2000. 橡胶延长因子的cDNA的克隆和序列分析. 华南热带农业大学学报, 2: 1. |
Adiwilaga K, Kush A. 1996. Cloning and characterization of cDNA encoding FDP synthase from rubber tree. Plant Mol Biol, 30: 935-946. DOI:10.1007/BF00020805 |
Amalon Z, Bangratz J, Chrestin H. 1992. Ethrel induced increases in the adenylated pool and transtonoplast pH with latex cells. Plant Physiol, 981: 270-1276. |
Asawatreratanakul K, Zhang Y W, Wititsuwannakul D, et al. 2003. Molecular cloning, expression and characterization of cDNA encoding cis-prenyltransferases from Hevea brasiliensis. Eur J Biochem, 270: 4671-4680. DOI:10.1046/j.1432-1033.2003.03863.x |
Broekeart W F, Lee H I, Chua N H, et al. 1990. Wound-induced accumulation of mRNA containing a hevein sequence in laticifers of rubber tree (Hevea brasi liensis). Proc Natl Acad Sci USA, 87: 7633-7637. DOI:10.1073/pnas.87.19.7633 |
Cardosa M J, Hamid S, Sunderasan E, et al. 1994. B-serum is highly immunogenic when compared to C-serum using enzyme immunoasssys. J Nat Rubber Res, 9: 205-211. |
Chye M, Kush A, Tan C T. 1991. Characterization of cDNA and genomic clones encoding HMG-CoA reductase from Hevea brasiliensis. Plant Mol Biol, 16: 567-577. DOI:10.1007/BF00023422 |
Chye M, Tan C T, Chua N H. 1992. Three genes encode HMG-CoA reductase in Hevea brasiliensis, hmg1 and hmg3 are differentially expressed. Plant Mol Biol, 19: 473-484. DOI:10.1007/BF00023395 |
D'Auzac J, Jacob J L, Prevot J C, et al. 1997. The regulation of cis-polyisoprene production. Present Res Plant Physiol, 1: 273-331. |
D'Auzac J, Prevot J C, Jacob J L. 1995. What' s new about lutoids?A vacuolar system model from Hevea latex. Plant Physiol Biochem, 33: 765-768. |
DennisM S, Henzel W J, Bell J, et al. 1989. Amino acid sequence of rubber elongation factor protein associated with rubber particles in Hevea latex. J Biol Chem, 264(31): 18618-18626. |
Gidrol X, Chrestin H, Tan H L, et al. 1994. Hevein, a lectin-like protein from Hevea brasiliensis(rubber tree)is involved in the coagulation of latex. J Biol Chem, 269: 9278-9283. |
Goyvaerts E, Dennis M, Light D, et al. 1991. Cloning and sequencing of the cDNA encoding the rubber elongation factor of Hevea brasiliensis. Plant Physiol, 97: 317-321. DOI:10.1104/pp.97.1.317 |
Han K H, Shin D H, Yang J M, et al. 2000. Genes expressed in the latex of Hevea brasiliensis. Tree Physiology, 20: 503-510. DOI:10.1093/treephys/20.8.503 |
Hao B Z, Wu J L. 2000. Laticifer differentiation in Hevea brasiliensis, induction by exogenous jasmonic acid and linolenic acid. Ann Bot, 85: 37-43. |
Jae-Heung Ko, Keng-See Chow, Kyung-Hwan Han. 2003. Transcriptome analysis reveals novel features of the molecular events occurring in the laticifers of Heveabrasiliensis (para rubber tree)Plant. Molecular Biology, 53: 479-492. DOI:10.1023/B:PLAN.0000019119.66643.5d |
Kush A, Goyvaerts E, Chye M L, et al. 1990. Laticifer specific gene express in Hevea brasiliensis. Proc Natl Acad Sci USA, 87: 1787-1790. DOI:10.1073/pnas.87.5.1787 |
Light D R, Dennis M S. 1989. Rubber elongation factor from Hevea brasiliensis. J Biol Chem, 264(31): 18608-18617. |
Miao Z, Gaynor J J. 1993. Molecular cloning, characterization and expression of Mn-SOD from the rubber tree. Plant Mol Biol, 23(2): 267-277. DOI:10.1007/BF00029003 |
Oh S K, Kang H, Shin D h, et al. 1999. Isolation, characterization and functional analysis of a novel cDNA clone encoding a small rubber particle protein from Hevea brasiliensis. J Biol Chem, 274(24): 17132-17138. DOI:10.1074/jbc.274.24.17132 |
Pujade-Renaud V, Clément A, Perrot-Rechenmann C, et al. 1994. Ethylene-induced increase in glutamine synthetase activity and mRNA levels in Hevea brasiliensis latex cells. Plant Physiology, 105: 127-132. DOI:10.1104/pp.105.1.127 |
Sivasubramaniam S, Vanniasingham V M, Tan C T, et al.1995.Characterization of HEVER, a novel stress-induced gene from Hevea brasiliensis, 29(1): 173-178 http://med.wanfangdata.com.cn/Paper/Detail/PeriodicalPaper_PM7579163
|
Takaya A, Zhang Y W, Asawatreratanakul K, et al. 2003. Cloning, expression and characterization of functional cDNA clone encoding geranylgeranyl diphosphatesynthase of Hevea brasiliensis. Biochemica et Biophysica ACTA, 16252: 14-220. |
Wu J L, Hao B Z, Tan H Y. 2002. Wound-induced differentiation in Hevea brasiliensis shoots mediated by Jasmonic acid. J Rubb Res, 5(1): 53-63. |