 2006, Vol. 42
2006, Vol. 42文章信息
- 杨敏生, 李志兰, 王颖, 王进茂, 梁海永.
- Yang Minsheng, Li Zhilan, Wang Ying, Wang Jinmao, Liang Haiyong.
- 双抗虫基因对三倍体毛白杨的转化和抗虫性表达
- Transformation and Expression of Two Insect-Resistant Genes to Hybrid Triploid of Chinese White Poplar
- 林业科学, 2006, 42(9): 61-68.
- Scientia Silvae Sinicae, 2006, 42(9): 61-68.
-
文章历史
- 收稿日期:2004-11-20
-
作者相关文章


2. 中国林业科学研究院亚热带林业研究所 富阳 311400;
3. 中国热带农业科学院热带生物技术研究所 热带作物生物技术国家重点实验室 海口 571101
2. Subtropical Forestry Institute, CAF Fuyang 311400;
3. Institue of Tropical Bioscience and Biotechnology, State Key Laboratory of Tropical Crops Biotechnology, Chinese Academy of Tropical Agricultural Sciences Haikou 571101
杨树是中国东北、华北、西北广泛栽种的主要速生造林树种之一, 也是世界上重要造林树种。近年来, 随着杨树人工造林面积的不断扩大, 杨树的虫害问题越来越突出。由于害虫危害, 大量的杨树生长不良, 丧失绿化、美化和防护效果, 甚至大面积死亡。因此, 培育抗虫杨树新品种已成为当务之急的重要任务。随着基因工程技术的发展, 植物抗虫基因工程育种受到世界各国的高度重视。国内外已有在杨树(Populus spp.)、落叶松(Larix spp.)、白云杉(Picea glauca)、枫香树(Liquidambar formosana)等转入抗虫基因的报道, 特别是杨树转基因研究取得了较大进展(王瑶等, 1999; 李志兰等, 2002)。1988年美国依阿华大学林学系McNabb等利用根癌农杆菌的Ti质粒为载体, 将马铃薯胰蛋白酶抑制剂(Pin-Ⅱ)基因成功地导入杨树NC5339无性系, 转化获得的抗虫杨已进入田间抗虫试验阶段。我国也在1993年获得了一批转BtCry1Ac基因的欧洲黑杨(P. nigra), 对舞毒蛾(Lymantria dispar)和杨尺蠖(Apocheima cinerarius)毒杀死亡率可达到80%~90%(田颖川等, 1993)。田颖川等(2000)和郑均宝等(2000)对BtCry1Ac基因部分片断的核苷酸序列进行了改造与慈菇蛋白酶抑制剂(API)基因构建了具有不同杀虫机制的双抗虫基因载体, 并成功的转化了白杨杂种——741毛白杨。在杨树方面进行转基因研究的还有McCown等(1991)、Klopenstein等(1993)、Howe等(1994)、Leple等(1992)、Confalonieri等(1998)、郝贵霞等(1999)、饶红宇等(2000)、杨敏生等(2005)。这些研究均获得了转化植株, 并表达出一定的抗虫性。
三倍体毛白杨是我国目前应用广泛的纸浆林工业用材树种(朱之悌, 1995)。为提高其抗虫性, 笔者将部分改造的Bt基因和慈菇蛋白酶抑制剂(API)基因构建在一个植物表达载体上, 通过根癌农杆菌介导将此表达载体上的双基因转入三倍体毛白杨中, 获得了一批对杨扇舟蛾等鳞翅目害虫具高抗性的植株。
1 材料与方法 1.1 材料 1.1.1 菌种和质粒含部分改造的BtCry1Ac基因(简称Bt基因)及慈姑蛋白酶抑制剂(API)基因的植物表达质粒载体为pBtiA, 所含的抗性筛选标记基因为卡那霉素抗性基因Npt Ⅱ(新霉素磷酸转移酶基因)。土壤农杆菌LBA4404及以上载体由中国科学院微生物研究所构建并提供(田颖川等, 2000)。
1.1.2 植物材料采用北京林业大学培育的三倍体毛白杨无性系73作为试验植物材料。
1.1.3 测试昆虫测试昆虫为杨扇舟蛾(Clostera anachoreta)和舞毒蛾。杨扇舟蛾新鲜卵块采自深州市北四王庄村苗圃, 舞毒蛾的卵块每年春季采自秦皇岛市。室内孵化幼虫, 人工饲养, 用于害虫检测。
1.2 方法 1.2.1 双抗虫基因的构建和改造将部分改造后的BtCry1Ac杀虫蛋白基因和慈菇蛋白酶抑制剂(API-A)基因构建在一个植物表达载体pBtiA上(田颖川等, 2000)。对BtCyr1Ac基因的两端进行改造, 在1—273 bp这个区域共改变了12个碱基, 涉及到11个密码子的改变, 一般是将第3位的AT改变为GC。还改变了2个潜在的mRNA转录终止信号AATTAA和AACCAA。这一区域对提高基因表达是非常重要的。在1 342—1 836区域通过9对引物共改变了63个碱基, 涉及60个密码子的改变, 同时改变了3个可能引起mRNA不稳定的ATTTA基序和8个AT富集区序。
1.2.2 双抗虫基因对三倍体毛白杨的转化诱导叶片不定芽分化和诱导芽增殖培养基为MS+6-BA0.6 mg·L-1+NAA0.1 mg·L-1, 诱导嫩茎生根培养基为1/2MS+IBA0.4 mg·L-1。采用农杆菌介导法进行基因转化, 卡那霉素筛选培养基附加50 mg·L-1卡那霉素, 300 mg·L-1羧苄青霉素。选择生根培养基附加50 mg·L-1卡那霉素和羧苄青霉素200 mg·L-1。
1.2.3 转化植株的分子生物学检测杨树DNA提取采用CTAB法进行。BtCry1Ac基因引物序列: 5′ CTGACGTAAGGATGACGCAC 3′, 5′ ACTATTGATAGTCGCGGCATC 3′。PCR扩增条件: 94 ℃50 s, 55 ℃60 s, 72 ℃100 s, 共30个循环。API基因引物序列: 5′ GCTGAATTCGACCATGGCGGCCTCCAACGCT 3′, 5′CGATGCCCAGCAAGGTTTT 3′。PCR扩增条件: 94 ℃50 s, 57 ℃50 s, 72 ℃50 s, 共30个循环。
采用地高辛试剂盒进行Southern blot检测, 试剂盒购自美国Roche Diagnostics公司。用BamHI对所提取DNA进行酶切, 置于37 ℃恒温箱中过夜。酶切产物进行琼脂糖凝胶电泳后, Southern吸印转至尼龙膜上, 以Digoxigenin标记的BtCry1Ac基因片段为探针, 与转至尼龙膜上PCR产物进行Southern杂交, 杂交后洗膜, 检测杂交信号。
蛋白质的提取按照Perlak等(1991)所述的方法进行。在ELSA分析中, 首先用1:2 500稀释的Bt蛋白鸡抗4 ℃包被过夜, 然后加已知浓度的标准抗原或植物蛋白溶液37 ℃反应3 h, 再加入1:1 000稀释的Bt蛋白鼠抗37 ℃反应2 h, 最后加入羊抗鼠酶标二抗37 ℃反应2 h, 每步均用PST洗板3次。最后加入底物避光反应30 min, 用BioRad550型酶标仪测定结果。ELISA试剂盒购自美国Agdia公司。
1.2.4 转基因植株的饲虫试验饲虫材料为苗圃试验地上的转基因和未转基因(CK)的三倍体毛白杨各株系1年生苗木新鲜叶片, 进行室内饲养。饲养瓶为高12 cm直径8.5 cm的罐头瓶, 用塑料薄膜扎紧瓶口, 细针扎孔通气。选取孵化1~2 d的1龄杨扇舟蛾和舞毒蛾开始饲养, 每瓶30头幼虫, 每2 d换1次新鲜叶片, 直到化蛹为止, 记载昆虫死亡数。试验结束时昆虫死亡数占参试总昆虫数百分比为害虫总死亡率。饲养21 d时对各系号的幼虫质量、平均体长和虫粪进行称量。
总死亡率=饲养末期的死亡总数/初期的饲养总数×100%;死亡指数=(∑天数×当天死亡数)/(总死亡数×试验总天数)。
2 结果与分析 2.1 三倍体毛白杨叶片再生体系的建立要确定完整而有效的转化系统, 分化和生根培养基的选择至关重要。通过对多个培养基比较分析, 培养基MS+6-BA 0.6 mg·L-1+NAA 0.1 mg·L-1附加15 g·L-1蔗糖和6 g·L-1琼脂较适合三倍体毛白杨叶片再生培养, 诱导率达到88.7%, 继代40 d左右平均芽高可达到1.2 cm, 且诱导出的芽健壮。在生根培养中, 采用1/2MS+IBA0.3 mg·L-1效果较好。接种后3 d就有生根现象, 10 d时生根率达到88.3%, 15 d后全部生根, 每个嫩茎小植株的主侧根数为8条, 平均根长达到3.3 cm。
2.2 农杆菌介导的基因转化以试管内无菌叶片为外植体, 采用农杆菌介导法进行基因转化。由于所用菌种带有抗卡那霉素基因(NPT Ⅱ, 新霉素磷酸转移酶标志基因), 因此在诱导和生根培养基中加入一定卡那霉素, 进行转基因植株的初步筛选。叶片经过浸菌和共培养后, 放置于附加50 mg·L-1卡那霉素, 300 mg·L-1羧苄青霉素的诱导不定芽分化的MS培养基中, 大约培养20~30 d, 叶片开始大部分坏死, 呈水渍状但在切口部位及叶柄剪口处出现米粒状愈伤组织, 经进一步培养, 诱导出不定芽, 约40~50 d形成丛状芽丛。能够诱导产生不定芽的叶片仅占培养叶片总数的18%左右。
在含有卡那霉素50 mg·L-1的生根培养基上对转基因植株作进一步筛选。结果表明:不同株系的三倍体毛白杨生根率和生根天数明显不同。最快的5 d即可生根, 最慢的16 d生根。生根率高的可以达到100%, 低的仅为48.4%。两周后统计根的生长状况, 最长的根长为13 mm, 最短的仅为1 mm。同时发现, 生根快的植株茎上部分生长也很快, 而且植株健壮。生根慢的植株前期常有蹲苗现象, 生长缓慢。所以, 一般根据嫩茎小植株在50 mg·L-1卡那酶素培养基上生根状况, 即可初步断定基因的转化成功与否。对照未转基因三倍体毛白杨在含50 mg·L-1卡那霉素的生根培养基中不能生根, 大部分植株黄化死亡。
2.3 转基因植株的DNA水平检测 2.3.1 BtCry1Ac基因和API基因的PCR检测提取抗卡那霉素株系基因组DNA, 以质粒pBtiA作阳性对照, 以未转基因三倍体毛白杨作为阴性对照, 进行PCR扩增。扩增产物的电泳图如图 1所示。


|
图 2 转基因再生三倍体毛白杨PCR检测API基因结果 Fig. 2 PCR detection of API gene of the transgenic hybrid triploid of Chinese White Poplar |
电泳分析表明:转双抗虫基因的三倍体毛白杨植株经PCR扩增, 各转基因植株均得到一条与阳性质粒作模板PCR扩增带大小相同的749 bp的条带, 而未转化三倍体毛白杨基因组未出现PCR扩增特异条带, 初步证明外源Bt基因已整合到杨树基因组中。
对蛋白酶抑制剂基因的PCR检测结果表明, 抗性植株均得到一条与阳性质粒作模板PCR扩增带大小相同的475 bp的条带, 而未转化三倍体毛白杨基因组未出现PCR扩增特异条带, 初步证明外源蛋白酶抑制剂基因已整合到杨树基因组中。
经过含卡那霉素培养基的生根及叶片再生多次筛选后得到的转化三倍体毛白杨株系, PCR检测表明80%的植株呈现阳性反应, 即在参加检测的36个转基因株系中, 28个株系PCR呈阳性反应。
2.3.2 BtCry1Ac基因的Southern Blot检测Southern杂交结果如图 3所示。转基因三倍体毛白杨各系号基因组DNA的BamHI酶切产物与Bt基因的探针杂交可产生一条大于2.3 kb的杂交带, 其中10号所产生的杂交带分子量较接近2.3 kb, 而11号、21号、7号所产生的杂交带分子量在5.0 kb, 未转基因植株没有产生任何杂交带。这一结果说明Bt基因已整合到三倍体毛白杨基因组DNA中, 由于采用单酶切, 每个转基因株系只出现1条杂交带, 初步证明是单拷贝整合。

|
图 3 转基因三倍体毛白杨基因组DNA的Southern杂交结果 Fig. 3 Hybridization fragment of genomic DNA of the transgenic hybrid triploid of Chinese White Poplar by Southern blotting |
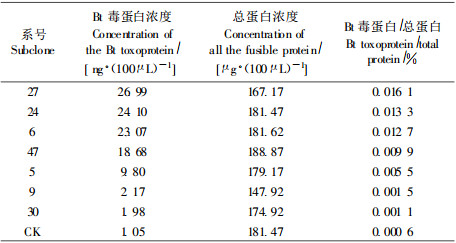
用抗Bt杀虫蛋白N端68 ku的抗血清对部分转基因杨树叶片蛋白提取物进行ELISA免疫测定。测定结果证明(表 1), 在7个转基因株系中有均有Bt杀虫蛋白的合成, 毒蛋白表达量为0.001 1%~0.016 1%, 均显著高于对照。未转基因的对照毒蛋白表达量仅为0.000 6%。
|
|
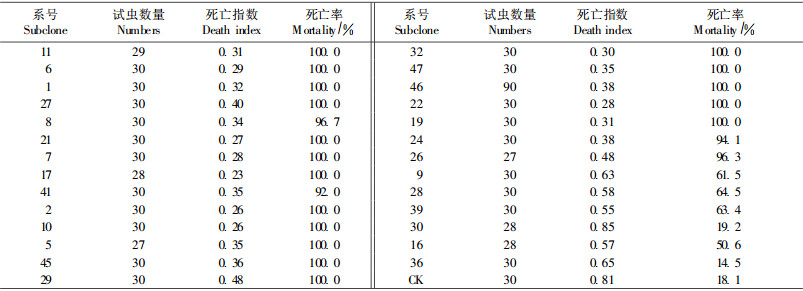
用不同系号的叶片饲养舞毒蛾幼虫, 以未转基因三倍体毛白杨作对照, 饲养21 d时幼虫死亡率和死亡指数列于表 2。从表 2可以看出:不同系号转基因三倍体毛白杨对舞毒蛾幼虫均有毒杀作用, 但死亡率不同。在处理21天时, 取食对照的幼虫死亡率为18.1%, 有21个系号幼虫死亡率在90%以上, 有4个系号幼虫死亡率为50%~65%, 30和36号株系的幼虫死亡率仅为19.2%和14.5%, 与对照未表现出明显差异, 基本未表现出抗虫性。死亡指数是幼虫死亡率和死亡速度的综合反映, 死亡指数越小, 表明前期死亡比例越大, 毒力越大; 死亡指数越大, 后期死亡比例越大, 毒力越小。死亡指数与死亡率紧密相关, 相关系数达到-0.915, 说明幼虫死亡率高的系号, 其死亡速度也较快。
|
|
用不同系号叶片饲养杨扇舟蛾21 d, 观察结果表明列于表 3。结果表明:不同系号转基因三倍体毛白杨对杨扇舟蛾均有致死效应, 但不同系号对幼虫的抗性不同。在参加测试的28个株系中, 有11个株系对三倍体毛白杨具有较强的抗虫性, 杨扇舟蛾幼虫死亡率在80%~ 100%之间, 死亡指数较小, 介于0.53~0.67之间; 有7个株系抗性中等, 幼虫死亡率在60%~80%之间, 死亡指数在0.48~0.75之间; 有10个系号抗虫性低, 幼虫死亡率在0~60%之间, 死亡指数大都介于0.70~0.90之间。未转基因三倍体毛白杨叶片喂养的昆虫幼虫死亡率为7.4%, 且死亡指数较大为0.77。
|
|
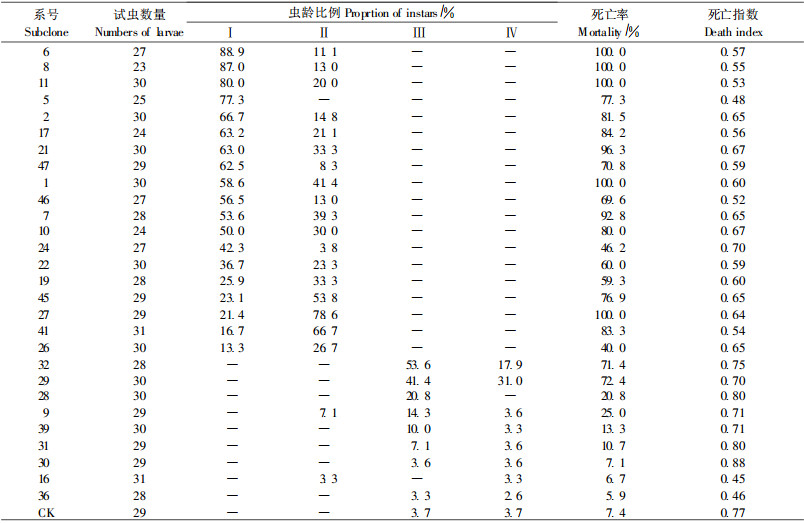
对杨扇舟蛾不同龄期死亡数进行调查, 计算各龄期死亡率(表 3)。结果表明:同一系号不同龄期的死亡率不同, 一般随着虫龄的增加幼虫死亡率降低。未转基因三倍体毛白杨有较低的死亡率, 且主要集中在3龄和4龄, 死亡率分别为3.7%和3.7%; 28个转基因株系中, 有12个株系对杨扇舟蛾1龄幼虫的致死率在50%以上, 而对2龄幼虫的致死率明显降低, 大部分均在40%以下, 这12个系号的幼虫大多都死于1龄和2龄, 进入3龄的极少。有7个株系的1龄幼虫致死率在10%~50%之间, 且大部分株系饲养的幼虫在2龄时死亡率高于1龄时的死亡率。未死亡且进入3龄和4龄的幼虫, 体质较弱, 且生长受到明显抑制, 与同龄对照相比虫体偏小; 有9个株系除29号和32号幼虫致死率较高外, 其它系号幼虫致死率均较低, 且绝大部分死于3龄和4龄。由此可见, 高抗系号对杨扇舟蛾1、2龄幼虫有很好毒杀效果, 进入3龄的机率很小; 中、低抗系号对杨扇舟蛾幼虫的毒杀效果依次减弱, 随着虫龄的升高幼虫本身抗性增强, 死亡率相对较低。
2.5.3 转基因株系对杨扇舟蛾幼虫发育的影响在用转基因三倍体毛白杨饲养的第21天时, 有22个株系仍有幼虫存活, 对存活的幼虫, 测量其体重、体长及虫粪重, 计算平均值, 各系号的比较结果列于表 4。
|
|
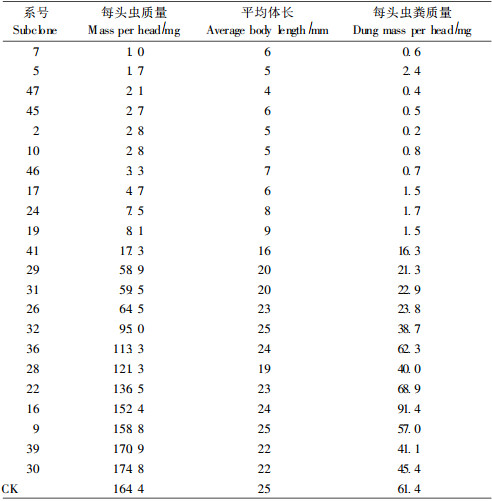
用转基因植株叶片饲养的杨扇舟蛾第21天时, 11个转基因株系饲养的幼虫单体质量均在20 mg以下, 绝大部分平均体长10 mm以下, 虫粪质量在每头2.5 mg以下, 说明这11个系号饲养的幼虫虫体较小, 代谢能力差, 且活虫数较少, 这些系号中多数为高抗或中抗株系。其余9个株系饲养的幼虫单体质量在55~160 mg之间, 平均体长在19~25 mm之间, 每头虫粪质量为20~95 mg, 说明这9个系号饲养的幼虫虫体较大, 代谢能力相对较强, 而且活虫数较多, 这些系号大多数为低抗株系。
3 讨论基因转化的效率与组织的培养效率和基因的转化系统相关, 筛选方法也是其中很重要的一环。为促进转化细胞的分裂和生长, 抑制非转化细胞, 须对培养的细胞施加选择压。由于农杆菌介导法的转化率多在10%以下。转化率低的原因之一是卡那霉素对转化细胞具有强烈的毒害作用, 致使大量的受体于选择培养基培养约一周时便死亡, 从而降低了转化细胞的再生率。为能减轻卡那霉素对芽再生的阻碍作用, 获得更多的转化芽, 很多研究者都采用推迟筛选的方法(Metz et al., 1995; Narasimhulu et al., 1996)。郝贵霞等(1999)认为:虽然推迟选择获得再生芽总数大大提高, 但获得真正的转化芽的频率没有显著提高, 并且对大量假阳性芽的筛选工作量巨大。所以又提出了分级分步筛选, 既共培养后先在低浓度的Kan选择培养基上诱导分化, 然后再移至含高浓度Kan的培养基上进行筛选。但这种选择方法较繁琐, 试验所需时间长, 还易使非转化细胞对选择会产生耐性, 增加假转化体和嵌合体出现的频率。本试验结果表明:叶片在共培养2 d后即用50 mg·L-1 Kan进行持续选择, 获得抗性芽后, 直接在含有50 mg·L-1 Kan生根培养基上进行生根培养。生根过程比诱导不定芽的过程对卡那霉素更加敏感, 未转基因的不定芽在卡那霉素含量为10 mg·L-1培养基上即不能生根。在含卡那霉素生根培养基上快速生根的不定芽, 再进行分化培养, 形成不同株系。采用这种两步筛选的方法, 不仅简单、节约了试验时间, 而且试验最终的转化率也较高, 最终诱导再生率为18%左右。
很多文献中提出了抗性植株PCR阳性率低的问题。据张艳贞等(2002)报道, 用PCR方法检测其转化得到的转化体113株, 获得PCR阳性植株仅为14株。她认为可能是由于所选用的抗性芽筛选方法不太合理, 导致PCR阳性率低。本研究采用卡那霉素进行诱导芽和生根筛选, 得到的抗性株系, 移植到苗圃中生长1年后, 进行PCR检测。在检测的42个株系中, 33个株系为阳性, 阳性率达到80%。
PCR检测容易出现假阳性, 可能是由于操作要求较低, 造成样本之间的交叉感染。而Southern杂交具有灵敏性高(可检出10~12, 即1 pg DNA样品), 特异性强(可鉴别出20个碱基对左右的同源序列)的特点, 是当前鉴定基因整合的权威方法。为了进一步证实PCR检测结果的准确性, 对转基因三倍体毛白杨4个高抗系号进行了Southern杂交。选用地高辛试剂盒进行Southern杂交, 证明测试的4个株系外源基因均以单拷贝的形式整合到三倍体毛白杨基因组中。
外源基因片段在植物DNA中插入位置及各种原因引起外源基因失活会导致转基因植物表现出不同的抗虫性, 因此转基因植物的目的基因的高效表达和稳定遗传尚不尽如人意。所以经PCR等分子生物学检测证明的转基因植株, 还需经过抗虫性生物测定试验以观察其实际抗性。对转基因植株进行毒蛋白表达量测定及进行实际饲虫试验, 是筛选具高抗虫性株系的必要环节。本研究用经过分子生物学检测证明已转入Bt-API基因的三倍体毛白杨株系1年生叶片进行了初步饲虫试验, 证明转基因株系对舞毒蛾和杨扇舟蛾均表现出了杀虫效果, 不同株系对幼虫的致死率不同。在参试的28个株系中, 有11个株系的植株叶片被舞毒蛾和杨扇舟蛾取食后, 幼虫死亡率均在80%以上, 占总虫试系号的39.3%;有7个株系对2种害虫的幼虫死亡率均在60%~80%之间, 占总虫试系号的25.0%;另有10个株系幼虫死亡率在50%以下, 有些株系基本未表现出抗虫性。这和其他文献报道一致。田颖川等(1993)年用含Bt基因的农杆菌LBA4404转化欧洲黑杨, 共获得54株转基因植株。15%的转基因植株对杨尺蠖的校正死亡率为80%~96%, 部分植株对舞毒蛾5~9天内的校正死亡率高达100%, 存活昆虫的生长和发育也明显受到抑制。Leple等(1995)发现一种半胱氨酸蛋白酶抑制剂(OC1)能抑制山杨叶甲(Chrysomela tremulae)的生长。用含OC1基因的农杆菌转化欧洲黑杨×美洲黑杨(P. deltoides), 用转基因植株饲喂山杨叶甲25天后, 幼虫死亡率为43.5%, 而对照只有4.5%, 同时对幼虫的生长和发育也有抑制作用。本研究获得的转基因三倍体毛白杨由于转入的是双价抗虫基因BtCry1Ac和慈菇蛋白酶抑制剂(API-A)基因, 因此转基因无性系除表现出杀虫活性外, 还应表现出抑制昆虫生长和发育的作用, 抗虫谱相对较广, 而且害虫很难对其产生耐受性, 因此, 具有更为稳定的抗虫效果。
研究结果表明:转基因植株不能将害虫全部杀死, 而且随着害虫的发育, 杀虫效果降低, 害虫在幼龄对转基因叶片最敏感, 死亡率较高。但随着虫体的发育, 死亡率降低, 说明害虫的抗性逐渐提高。转基因植株对害虫的发育具有较大的影响, 可以明显降低害虫的体质量和体长, 说明虽然没有致死, 但严重影响其生长发育, 有些个体不能完成其发育历期, 不能正常化蛹, 繁衍后代, 这可能是双基因共同作用的结果。文献报道, Bt基因对昆虫的生物学如生活、生长、发育和繁殖均发生显著影响, 常表现为幼虫存活率、化蛹率及蛹羽化率降低, 幼虫质量、蛹质量变小, 幼虫发育减慢, 成虫产卵率降低, 严重的导致死亡; 蛋白酶抑制剂基因可明显抑制幼虫生长并推迟化蛹, 干扰昆虫蜕皮过程和免疫功能, 导致昆虫不能正常发育(俞志华, 2000)。由于741杨中转入的是双价抗虫基因Bt和慈姑蛋白酶抑制剂(API)基因, 因此转基因株系除表现出杀虫活性外, 还应表现出抑制昆虫生长和发育的作用。但目前我们还不能证明这2个基因在植物中都得到了有效表达。
用美国Agdia公司生产的Cry1Ab/Cry1Ac试剂盒, 对初步虫试筛选出的8个株系进行了ELISA分析, 测定结果与虫试结果基本一致, 相关系数达到0.827的极显著水平, 说明2种检测方法都比较可靠。2种方法结合, 可以准确、快速地对大量转化植株进行抗虫性筛选。
转基因植株的应用可以巨大的抗虫效果, 但其缺点也是明显的。除了环境安全上的问题外, 害虫的耐受性也是一个不容忽视的问题。随着转抗虫基因杨树的田间释放, Bt的局限性正逐渐显示出来, 表现在杀虫谱带窄、毒力不够强, 更为令人关心的是, 广泛采用这种转抗虫基因杨树后是否会使害虫产生抗性演化。由于树木长期的生长过程中基因始终不变, 而昆虫经过许多世代演化后, 一旦对这种转基因树木产生抗性, 那么转基因植株将失去其自身的价值。对此, 许多学者指出要将昆虫的抗性演化控制在一定程度。采用多种基因转化植物、将不同基因转化的植物混栽、工程植株和非工程植株混栽等技术措施, 这些都将大大减缓昆虫的抗性演化, 这是需要在今后进一步研究的工作。
郝贵霞, 朱祯, 朱之悌. 1999. 豇豆蛋白酶抑制剂基因转化毛白杨的研究. 植物学报, 41(12): 1276-1282. DOI:10.3321/j.issn:1672-9072.1999.12.005 |
李志兰, 杨敏生. 2002. 杨树基因工程育种研究进展. 河北农业大学学报, 25(Sup.): 145-148. |
饶红宇, 伍宁丰, 陈英, 等. 2000. 杨树NL-80106转Bt基因植株的获得及抗虫性. 植物资源与环境学报, 9(2): 1-5. DOI:10.3969/j.issn.1674-7895.2000.02.001 |
田颖川, 韩一凡. 1993. 抗虫转基因欧洲黑杨的培育. 生物工程学报, 9(4): 291-297. DOI:10.3321/j.issn:1000-3061.1993.04.017 |
田颖川, 郑均宝, 虞红梅, 等. 2000. 转双抗虫基因杂种741毛白杨的研究. 植物学报, 42(3): 263-268. |
王瑶, 林木兰, 沈锡辉, 等. 1999. 农杆菌介导的木本植物遗传转化. 生物技术通报, 6: 23-27. |
杨敏生, 高宝嘉, 王进茂. 2005. 转双抗虫基因741杨基本特性分析. 林业科学, 41(1): 91-97. |
俞志华. 2000. 抗虫基因的抗虫原理及其应用现状和展望. 生物学通报, 35(7): 8-10. DOI:10.3969/j.issn.0006-3193.2000.07.003 |
郑均宝, 梁海永, 田颖川, 等. 2000. 转双抗虫基因741毛白杨的选择及抗虫性. 林业科学, 36(2): 55-62. DOI:10.3969/j.issn.1672-8246.2000.02.014 |
朱之悌, 林惠斌, 康向阳. 1995. 毛白杨异源三倍体B301等无性系选育的研究. 林业科学, 31(6): 499-505. |
张艳贞, 王罡, 胡汉桥, 等. 2002. 农杆菌介导将Bt杀虫蛋白基因导入优良玉米自交系的研究. 遗传, 24(1): 35-39. |
Confalonieri M, Allegro G, Balestrazzi A. 1998. Regeneration of Populus nigra transgenic plants expressing a Kunitz proteinase inhibitor (Kti 3)gene. Mol Breeding, 4: 137-145. DOI:10.1023/A:1009640204314 |
Howe G T, Goldfarb B. 1994. Agrobacterium-mediated transformation of hybrid poplar suspension cultures and regeneration of transformed plants. Plant Cell, Tissue and Organ Culture, 36: 59-71. DOI:10.1007/BF00048316 |
Klopensten N B, McNabb S H, Hart E. 1993. Transformation of Populus hybrids to study and improve pest resistance. Silvae Genetica, 42(2-3): 86-90. |
Leple J C, Brasileiro A C M. 1992. Transgenic poplars: expression of chimeric genes using four different constructs. Plant Cell Reports, 11: 137-141. |
McCown B H, McCabe D E, Russell D R. 1991. Stable transformation of Populus and incorporation of pest resistance by electric discharge particle acceleration. Plant Cell Reports, 9: 590-594. |
McNabb H S. 1991. A field trial of transgenic hybrid poplar trees: establishment and growth through the second season. Agroculture Resarch Institute, 9: 155-159. |
Metz T D, Dixit R, Earle E D. 1995. Agrobacter iumtume faciens-mediated transformation of broccoli and cabbage. Plant Cell Reports, 15: 287-292. DOI:10.1007/BF00193738 |
Narasimhulu J, Hodges T K. 1996. Agrobacterium tumefaciens-mediated transformation of japonica and indica rice varieties. Planta, 6: 12-17. |
Perlak F J, Deaton R W, Armstrong T A, et al. 1990. Insect resistant cotton plants. Biotechnology, 8: 939-943. |