 2006, Vol. 42
2006, Vol. 42文章信息
- 吴家兵, 关德新, 施婷婷, 韩士杰, 于贵瑞, 孙晓敏.
- Wu Jiabing, Guan Dexin, Shi Tingting, Han Shijie, Yu Guirui, Sun Xiaomin.
- 非生长季长白山红松针阔叶混交林CO2通量特征
- Non-growing Seasons CO2 Flux of a Broad-leaved Korean Pine Forest in Changbai Mountain
- 林业科学, 2006, 42(9): 1-6.
- Scientia Silvae Sinicae, 2006, 42(9): 1-6.
-
文章历史
- 收稿日期:2005-04-28
-
作者相关文章



2. 中国科学院研究生院 北京 100049;
3. 中国科学院地理科学与资源研究所 北京 100101
2. Graduate school of Chinese Academy of Sciences Beijing 100049;
3. Institute of Geographic Sciences and Natural Research, Chinese Academy of Sciences Beijing 100101
森林是陆地生态系统的主体, 在生长过程中吸收了大量的CO2, 并将其长期贮存(徐德应, 1996)。Potter等(1999)研究发现, 全球陆地生态系统中46%的碳贮存在森林中, 1990—1995年由森林生长形成的净碳吸收量达0.9 Pg·a-1。在全球气候变化的背景下, 对森林生态系统碳收支特征的研究已成为关注的热点(于贵瑞等, 2001)。
目前, 针对生长季森林碳交换特征的研究已有很多(关德新等, 2004; 于贵瑞等, 2004)。但在非生长季, 由于植被、凋落物及土壤微生物的呼吸作用, 森林通常表现为碳释放, 特别是冬季持续时间较长的寒带及部分温带森林, 它们在非生长季释放出大量的CO2, 很大程度上决定了全年的碳源和碳汇强度(Falge et al., 2002)。因此, 研究非生长季森林与大气间的CO2通量特征, 对准确评价森林生态系统的碳收支状况具有重要价值。但观测期间受雪、霜以及低温条件限制, 相关研究开展的很少。
20世纪80年代初, 涡度相关法开始应用到森林碳交换研究中(Verma et al., 1986)。目前已成为国际上森林-大气物质与能量交换研究的主流方法(Baldocchi et al., 2001)。近年来, 该方法引入我国(黄耀, 2002), 并以其高精度、高时间分辨率等优点被广泛地应用到森林、农田、草地等生态系统碳收支研究中。本项研究依托中国通量网(China FLUX), 对CO2通量及其他气象因子进行了长期、连续的观测, 旨在探明长白山红松针阔叶混交林这一典型东北森林生态系统在非生长季的碳交换特征, 其结果对评价我国北方森林碳源或碳汇强度具有重要参考价值。
1 研究地概况本研究在中国科学院长白山森林生态系统定位站1号标准地阔叶红松(Pinus koraiensis)林内进行(42°24′ N, 128°6′ E), 海拔738 m。观测场附近下垫面均质、平坦, 土壤为山地暗棕色森林土, 主要乔木为红松、紫椴(Tilia amurensis)、蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandshurica)、色木槭(Acer mono)等。林分为复层结构, 平均株高26 m, 立木株数约560株·hm-2。
2 研究方法 2.1 涡度通量观测观测场建有高62 m的微气象观测塔, 在观测塔40 m (1.5倍林冠高)高处的主风方向铰接一支臂, 安装三维超声风速仪(CSAT3, Campbell Inc., USA)测量垂直风速与空气虚温脉动。CO2与水汽浓度脉动采用开路式红外气体分析仪(Li7500, Li-Cor, USA)测量, 脉动信号采样频率为10 Hz。所有通量数据通过CR5000数据采集器(Campbell Inc., USA)采集。观测日期为2002年10月11日(阔叶树落叶)至2003年4月20日(植被开始萌芽)。
观测塔配备小气候梯度观测系统, 同步进行气象要素梯度观测。数据采样频率为2 Hz, CR10X-TD数据采集器(Campbell Inc., USA)每30 min自动记录平均风速、温度、气压、光合有效辐射等常规气象信息。另外, 观测塔还配备CO2浓度7层(2.5、8、22、26、32、50和60 m)廓线系统(Li820, Li-Cor, USA), 进行CO2浓度梯度观测。
2.2 数据处理 2.2.1 涡度通量计算CO2涡度通量通过垂直风速w与CO2浓度c脉动值的协方差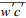
当红外气体分析仪实测的CO2或H2O脉动量是气体浓度而非混合比时, 为消除下垫面与大气间感热与潜热产生的密度脉动效应, 必须对涡度通量
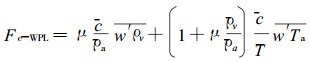
|
(1) |
式中:Fc-WPL为WPL修订项(mg·m-2s-1); c为CO2浓度(mg·m3); T为虚温(℃); μ为干燥空气和水气分子质量之比; 上横线表示单元数据平均; ρa与ρv分别为干燥空气与水气密度(mg·m-3), Ta为空气温度(℃)。
2.2.3 贮存项计算当大气层节稳定或湍流混合作用较弱时, 土壤与植被呼吸释放的CO2不能达到仪器测定高度, 在传感器以下形成积聚。在此采用Baldocchi等(1988)提供的简化方法计算探头以下气层储存项:
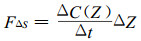
|
(2) |
式中:ΔC (Z)为观测高度处的CO2浓度变化(mg·m-3), Δt为采样周期(min), ΔZ为观测高度(m)。
2.2.4 数据插补受雨、雪或仪器标定、维护等影响, 涡度相关数据存在奇异值或缺测。对于奇异值, 目前国际通量网(FLUXNET)各个台站的判别标准与插补方法不一。本文根据相关文献(Falge et al., 2001)及CO2通量特征曲线分析, 确定CO2通量域值。剔除间隔奇异值后, 采用邻近值线性内插; 对于连续奇异值, 参考相似气象条件的邻近时段观测值进行插补; 若一天中奇异值超过40%, 则舍去该日全部信息, 根据CO2日交换量与土壤温度的回归关系计算日总量。
3 结果与分析 3.1 CO2浓度变化当森林与大气间CO2传输满足一维传输模式时, 林内CO2浓度的时空变化特征能够对森林碳收支动态提供定性描述。图 1所示的为林内7个观测高度CO2浓度的几何平均。总体来说, 在秋末(10月中下旬)与早春(4月上中旬)浓度较低, 冬季最高, 变动范围为366~411 μmol·mol-1, 最高值出现在1月。这一变化特征与下文中讨论的森林-大气CO2通量变化特征稍有差异, 估计与观测期间气压变化及大尺度环流有关。CO2浓度日振幅以秋末最大, 早春次之, 冬季最小。平均振幅为10 μmol·mol-1, 高值一般出现在凌晨6:00—7:00, 低值出现在午后2:00左右。除了日变化和季节变化, CO2浓度还存在约10天左右的中尺度天气过程造成的波动, 最大振幅可达30 μmol·mol-1。整个非生长季CO2平均浓度为389 μmol·mol-1, 较全年CO2浓度平均水平高出12 μmol·mol-1 (吴家兵等, 2005)。

|
图 1 林内CO2浓度季节变化 Fig. 1 Seasonal dynamics of CO2 concentration in forest |
图 2为10月12—13日7层廓线系统实测的CO2浓度在时间和空间上的演化图。可见:日出后, 各层CO2浓度持续降低, 如图 2中8:00—16:00所示, 在20~30 m高度处存在一个明显的浓度低值区, CO2呈现上层和下层逐渐向中间冠层浓度递减的趋势, 表明在冠层位置存在光合吸收现象。但在冠层以上(30~60 m), 由于湍流交换活跃, 浓度变化不大, 特别是白天, 浓度梯度接近于零。日落后, 林内各层CO2浓度逐渐升高, 以近地层浓度变化尤为显著, 表明期间CO2交换过程主要是土壤-大气的呼吸释放。林内各层CO2浓度最高值通常出现在凌晨, 即林内逆温层开始打破的日出前后。近地面层2.5 m处受下木层植被生理活动和大气条件日变化的影响, 也存在着明显的昼低夜高的日变化, 但由于土壤呼吸的决定作用, 全天的CO2浓度都相对较高。

|
图 2 林内CO2浓度时空演化图(10月12—13日) Fig. 2 Temporal-spatial evolution of CO2 concentration in forest (from October 12 to October 13) |
非生长季森林-大气CO2交换具有明显的季节变化趋势, CO2通量变动范围为-0.3~0.5 mg·m-2s-1, 秋末最大, 早春次之, 冬季较小且相对恒定。
秋末, 虽然正、负CO2通量均呈逐渐减弱趋势, 但森林-大气CO2交换能力依然较强(图 3)。森林夜间表现为强释放, 日间表现为一定的吸收。在晴朗天气里, 上午10:00左右, 森林大气CO2交换达到平衡, 森林生态系统转变为净吸收, CO2交换速率最大值约为-0.3 mg·m-2s-1, 出现在午间12:00~13:00;日落前2小时左右再次达到收支平衡, 森林转变为大气CO2的源; 夜间, CO2通量为0.1~0.3 mg·m-2s-1, 20:30~22:00交换速率最大, 峰值为0.5 mg·m-2s-1。从净光合和净呼吸通量的数量大小及持续时间来看, 森林在秋末以土壤、植被的呼吸通量为主。冬季, 森林-大气CO2通量较小, 且通量变化相对恒定, 通常小于0.1 mg·m-2s-1, 特别是1和2月, CO2通量仅为0.02~0.03 mg·m-2s-1, 而且此期间无明显的日变化。这可能与覆雪导致土壤温度基本上没有日变化有关; 也可能与积雪对土壤呼吸释放出的CO2存在缓释效应有关。4月上旬, 地表积雪融化, 5 cm土壤温度徘徊在0 ℃左右, 但森林却出现一个与温度不相称的高排放速率。至4月中旬, 开始出现负的通量, 并逐渐增强, 森林总体表现为由一定强度的碳源向碳汇转变。

|
图 3 阔叶红松林CO2通量日变化过程 Fig. 3 Daily course of CO2 flux in broad-leaved Korean Pine forest |
通常将5 ℃作为温带森林进行光合作用的生物学最低温度(贺庆堂, 1988)。从图 3可以看出, 10月下旬日平均温度已低于0 ℃, 但仍然出现负通量, 对期间日最高气温在5 ℃以下的观测数据(午间10:00—5:00)分析表明:CO2通量与光合有效辐射的相关性较显著(图 4)。由于林内阔叶树及冠下草本均已落叶, 据此推测, 常绿的红松在温度低于生物学最低温度以后, 仍可能存在微弱的光合吸收作用。由于负值持续时间较短, 且绝对值较小, 期间森林总体表现为微弱的碳源。

|
图 4 CO2通量与光合有效辐射的关系(10月20—30日) Fig. 4 Linear relationship of CO2 flux and photosynthetic active radiation (from October 20 to October 30) |
非生长季, 森林的主要生理活动由光合吸收转变为呼吸释放。很多文献报导过森林各组分(树干、土壤、凋落物)的呼吸强度主要受温度控制(刘建军等, 2003; 王淼等, 2005)。为此, 对空气温度和净生态系统碳交换量(net ecosystem carbon exchange, 简称NEE)以及土壤温度和NEE之间的相关性也分别进行了分析。根据空气温度和土壤温度对NEE变化的解释比例, 即相关系数R2判断, 5 cm土壤温度对NEE有更好的预测作用(图 5)。在5 cm土壤温度高于0 ℃时, NEE与日平均土壤温度指数相关, 相关系数R2为0.67, 达到显著水平(n=52, P < 0.05)。从图 5可以看出, 较多的有效数据集中在0 ℃附近, 且NEE波动较大, 特别是存在一些在较高温度才会出现的高NEE值。实测的CO2通量时间序列显示低值主要出现在秋末, 高值主要出现在早春融雪阶段。有学者报导在积雪融化时, 雪下及土壤中富集的CO2有一快速释放的过程(Goulden et al., 1997), 文中NEE低温高值现象可能正是这个原因所致。

|
图 5 净生态系统碳交换量与5 cm深土壤温度关系 Fig. 5 Relationship between net ecosystem carbon exchange and 5 cm depth soil temperature |
非生长季, 净生态系统碳交换值较大的过程主要集中在秋末与早春, 如图 6所示。秋末, 由于只在午间存在数小时的净吸收, 全天NEE表现为正值, 森林向大气排放CO2, 随着土壤温度降低, NEE随之降低; 冬季, 森林具有微弱但相对恒定的CO2排放, 特别是在1、2月, 日排放量多徘徊在0.4 g C·m-2左右; 初春(4月上旬), 覆雪融化, 土壤解冻, NEE出现1个正的峰值, 但随后显著下降, 在融雪阶段出现明显的CO2释放高峰, 说明积雪对CO2有一定的富集作用。另外, 从其变化曲线可以预测出, 在4月下旬或5月森林会由CO2的释放源转变为吸收汇。观测期间(190 d), 长白山红松针阔叶混交林净释碳交换量为127 g C·m-2, 整体表现为一定强度的碳释放。

|
图 6 非生长季净生态系统碳交换日总量 Fig. 6 Daily change of net ecosystem carbon exchange in non-growing seasons |
非生长季, 长白山红松针阔叶混交林CO2通量为-0.3~0.5 mg·m-2s-1, 释放的碳总量为127 g C·m-2, 整体表现为一定强度的碳释放。CO2通量具有明显的季节变化趋势。秋末, 正、负CO2通量均呈减弱趋势, NEE降低; 冬季, CO2排放速率相对恒定, 无明显的日变化; 初春, 正、负CO2通量均显著增大, 森林在融雪时候有1个CO2排放高峰。当土壤温度高于0 ℃时, 森林每天释放CO2的速率与5 cm深土壤温度呈指数相关。
在非生长季, 部分森林仍有一定的碳吸收能力, 甚至在冬季还有碳吸收(Dolman et al., 2002; Miyazawa et al., 2005)。长白山红松针阔叶混交林在温度低于生物学最低温度时仍有数小时表现为一定强度的CO2吸收。初步判断, 常绿红松在低温时可能仍维持着一定的光合能力。但由于野外观测的复杂性和涡度相关仪器系统在冬季低温条件下奇异观测值增多的干扰, 上述结论仍有待进一步的试验验证, 并从机理上探明长白山红松针阔叶混交林非生长季吸收CO2的物理和生理机制。
Li7500红外气体分析仪和CSAT3超声风速仪均可以在-20 ℃以下的环境温度中工作, 但试验中发现, 当空气温度低于-5 ℃时, 观测结果奇异值开始增加, 当温度低于-10 ℃时, 出现了大量的负通量(超过70%)。由于夜间森林碳吸收现象从物理学和生物学角度都无法解释, 因此目前认定这类观测数据不合理。文中为了估算长期的净碳交换量, 根据秋末和早春的数据与土壤温度间的关系对日总量做了简单插补, 虽然存在一定的不确定性, 但非生长季森林-大气间CO2交换主要集中在秋末和早春, 当空气温度降至-5 ℃时, 日净碳交换量通常小于0.5 g C·m-2, 对非生长季NEE总量估算影响不会很大。但这也提出了一个目前困扰着国内多个通量观测站的难题(于贵瑞等, 2004), 即冬季低温条件下, 大量奇异值, 特别是负值出现的原因究竟是什么?笔者根据对CO2浓度与三维风速实测时间序列的谱分布检验及平稳性检验初步认为, 仪器自身在低温时存在性能局限的可能性不大, 密度脉动效应可能是冬季通量观测出现负值的主要原因。
Web等(1980)针对涡度通量观测时感热与潜热传输会造成密度脉动效应, 提出了WPL修订理论, 并作为1个相对成熟的方案, 一直被学术界引用至今。但最近有学者提出, 在高风速、强湍流观测条件下, 需要对WPL修订项做压力产生的密度脉动的补充修订(Massman et al., 2002)。在冬季, 风速增加, 相应的压力脉动造成的通量损失会增加, 有可能造成负值的出现。但对于森林观测站, 这一建议却没有引起足够的重视, 加之压力脉动观测成本较高, 即使在国外, 这一补充修订做的也很少。根据式(1), 密度脉动修订量与观测层背景CO2浓度值正相关, 而与实际通量大小无关。因此, 对于CO2通量相对较小的非生长季森林, 修订比例通常较大, 个别时段修订量甚至超过实测值本身。在冬季低温、强风的特殊观测环境下, 修订方法的少许改进, 会大大改善最终结果。由于观测手段的局限性, 关于压力脉动项造成的冬季观测负值只是推测, 仍有待于进一步的试验验证。
关德新, 吴家兵, 于贵瑞, 等. 2004. 气象条件对长白山红松针阔叶混交林CO2通量的影响. 中国科学:D辑, 34(增刊Ⅱ): 103-108. |
黄耀. 2002. 中国陆地和近海生态系统碳收支研究. 中国科学院院刊, 17(2): 104-107. DOI:10.3969/j.issn.1000-3045.2002.02.006 |
贺庆棠. 1988. 气象学:修订版. 北京: 中国林业出版社, 47.
|
刘建军, 王得祥, 雷瑞德, 等. 2003. 秦岭天然油松、锐齿栎林地土壤呼吸与CO2释放. 林业科学, 39(2): 8-13. |
于贵瑞, 牛栋, 王秋凤. 2001. 《联合国气候变化框架公约》谈判中的焦点问题. 资源科学, 23(6): 10-16. DOI:10.3321/j.issn:1007-7588.2001.06.003 |
于贵瑞, 张雷明, 孙晓敏, 等. 2004. 亚洲区域陆地生态系统碳通量观测研究进展. 中国科学:D辑, 34(增刊Ⅱ): 15-29. |
王淼, 姬兰柱, 李秋荣, 等. 2005. 长白山地区红松树干呼吸的研究. 应用生态学报, 16(1): 7-13. DOI:10.3321/j.issn:1001-9332.2005.01.002 |
吴家兵, 关德新, 赵晓松, 等. 2005. 长白山红松针阔叶混交林CO2浓度特征. 应用生态学报, 16(1): 49-53. DOI:10.3321/j.issn:1001-9332.2005.01.010 |
徐德应. 1996. 中国大规模造林减少大气碳积累的潜力及其成本效益分析. 林业科学, 32(6): 491-499. |
Baldocchi D, Falge E, Gu L H, et al. 2001. FLUXNET: Anewtool to study the temporal and spatial variability of ecosystem-scale carbon dioxide, watervapor, and energyflux densities. Bulletin of the American Meteorological Society, 82(11): 2415-2434. DOI:10.1175/1520-0477(2001)082<2415:FANTTS>2.3.CO;2 |
Baldocchi D, Hicks B B, Meyers TP. 1988. Measuring biosphere-atmosphere exchanges of biologically related gases with micrometeorological methods. Ecology, 69: 1331-1340. DOI:10.2307/1941631 |
Dolman AJ, Moors EJ, Elbers J A. 2002. The carbon uptake of a midlatitude pineforest growing on sandy soil. Agricultural and Forest Meteorology, 111: 187-202. DOI:10.1016/S0168-1923(02)00022-9 |
Falge E, Baldocchia D, Olsonb R, et al. 2001. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agricultural and Forest Meteorology, 107: 43-69. DOI:10.1016/S0168-1923(00)00225-2 |
Falge E, Baldocchi D, Tenhunen J, et al. 2002. Seasonality of ecosystem respiration and gross primary production as derived from FLUXNET measurements. Agricultural and Forest Meteorology, 113: 53-74. DOI:10.1016/S0168-1923(02)00102-8 |
Goulden ML, Daube B C, Fan S M, et al. 1997. Physiological responses of a black spruce forest to weather. Journal of Geophysical Research, 102(D24): 28987-28996. DOI:10.1029/97JD01111 |
Massman WJ, Lee X H. 2002. Eddy covariance flux corrections and uncertainties inlong-termstudies of carbon and energy exchanges. Agricultural and ForestMeteorology, 113: 121-144. |
Miyazawa Y, Kikuzawa K. 2005. Winter photosynthesis by saplings of evergreen broad-leavedtrees in a deciduous temperate forest. NewPhytologist, 165(3): 857-866. |
Potter CS, Klooster S A. 1999. Detecting aterrestrial biosphere sinkfor carbon dioxide :Interannual ecosystem modeling for the mid-1980s. Climatic Change, 42: 489-503. DOI:10.1023/A:1005449017059 |
Verma S B, Baldocchi D D, Anderson E, et al. 1986. Eddyfluxes of CO2water vapor, andsensible heat over a deciduous forest. Boundary Layer Meteorology, 36: 71-91. DOI:10.1007/BF00117459 |
Web E K, Pearman GI, Leuning R. 1980. Correction of flux measurements for density effects dueto heat and water vapour transfer. The Quarterly Journal of theRoyal Meteorological Society, 106: 85-100. DOI:10.1002/qj.49710644707 |