 2006, Vol. 42
2006, Vol. 42文章信息
- 张飞萍, 陈清林, 施友文, 方妙辉, 尤民生.
- Zhang Feiping, Chen Qinglin, Shi Youwen, Fang Miaohui, You Minsheng.
- 毛竹林林下植被、竹叶化学成分与节肢动物群落的关系
- Relationships among Undergrowths, Contents of Chemical Substances in Leaves and Arthropod Community in Phyllostachys heterocycla cv. pubescens Forest
- 林业科学, 2006, 42(8): 50-56.
- Scientia Silvae Sinicae, 2006, 42(8): 50-56.
-
文章历史
- 收稿日期:2004-09-27
-
作者相关文章


2. 厦门大学生命科学院 厦门 361000;
3. 福建省三明市三元区林业局 三明 365000
2. Life Science College, Xiamen University Xiamen 361000;
3. Sanyuan Forestry Bureau of Sanming, Fujian Province Sanming 365000
栖境变化是人为干扰引起生态系统结构与功能变化的最主要原因(Diamond,1989),定量研究栖境与生物种群或群落的关系有助于揭示系统变化的过程与机理。毛竹(Phyllostachys heterocycla cv. pubescens)林是中国南方典型的人工林生态系统,在大面积竹木混交林纯林化后,对竹林的施肥、垦复、劈草和化学除草等管理措施已是除伐竹和挖笋外最常见的人为干扰,这些干扰均显著地改变了竹林下层的植被和土壤条件(张飞萍等,2004),对节肢动物群落的结构与组成产生了重要影响(张飞萍等,2005a)。显然,林下植被的改变将直接影响到以此为栖境的林下节肢动物的物种组成与时空分布格局,然而,由于林下与冠层在时间上的同步和空间上的相邻,在群落长期的演化进程中,2层次的节肢动物形成了密切的能量流动和物种交流等关系(张飞萍等,2005b ),因而林下植被的改变也影响着竹林冠层节肢动物的物种组成与分布。林下土壤条件如肥力、保水能力、透气性和疏松度等的改变则直接影响着毛竹的生长情况和生理生化状况,并因此而影响着植食性种群的时空动态,进而对整个群落产生影响。许多资料表明:施肥、灌溉等农业措施可改变植物物质组成及其质量,从而对植食性昆虫产生正面或负面影响,且一般以其营养和次生物质的变动影响昆虫的行为、生长发育和生殖(钦俊德,1980)。蛋白质和可溶性糖是植物中C、N的主要存在形式,其质和量足以影响昆虫的生命过程。植物体内某些次生代谢物质,如酚、单宁、类黄酮、生物碱和萜类等也可通过对植食者的毒害或对其食物利用率、消化力、取食量和嗜食性等的影响,导致植食性种群增长、生存和繁殖力的显著改变(Chan,1978;Schultz et al., 1982;Klocke et al., 1982;Berenbaum,1983;Waage et al., 1984;Hedin,1992)。本文从林下植被和毛竹生理的角度定量探讨栖境变化与节肢动物群落的关系,不仅为害虫成灾机理解释和生态控制提供重要的基础,还可为深入解析人为干扰引起生态系统结构与功能变化的过程与机理提供基础依据。
1 材料与方法 1.1 试验标准地概况和管理措施的划分与参考文献张飞萍等(2004)相同。
1.2 群落的调查与分析与参考文献张飞萍等(2005c)相同。
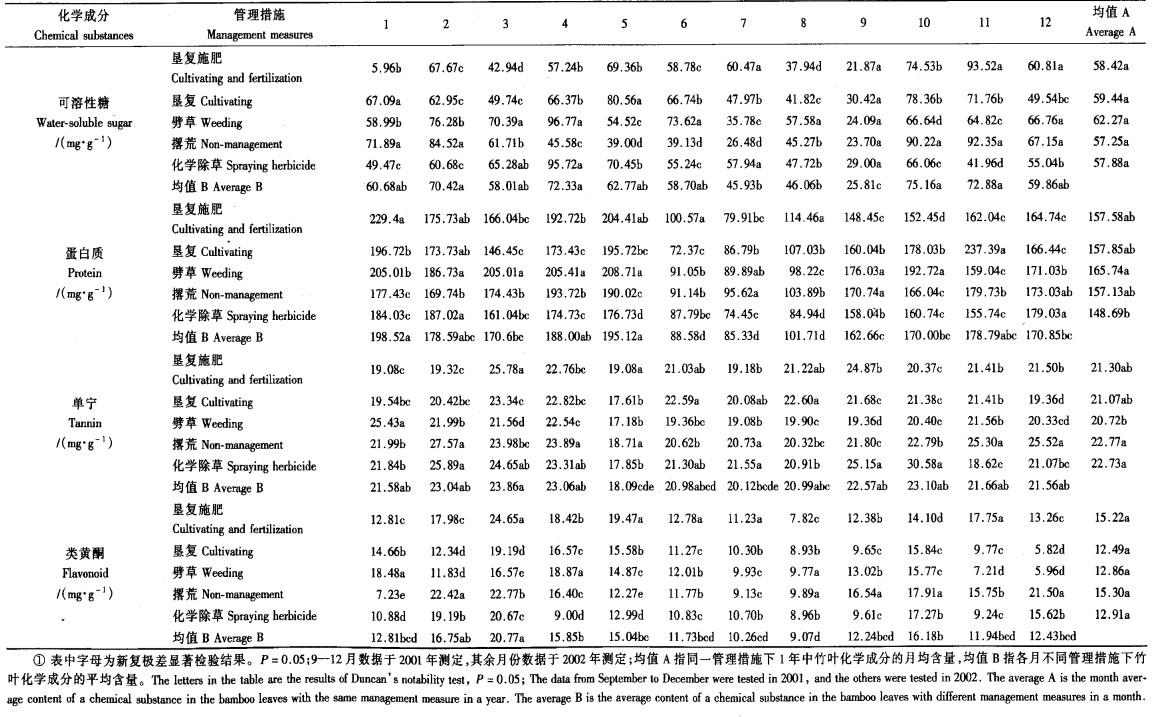
1.3 竹叶化学成分测定以5点取样法在1—6号标准地随机抽取20样株并标记,2001年9月—2002年8月,每月15日从各样株上随机剪取上、中、下部各1小枝,以标准地为单位带回室内,立即剪取叶柄以上部位叶片,用自来水洗净后置于80~100 ℃烘箱内烘干12 h,粉碎,过40目筛,混匀,贮于磨口瓶中待测。可溶性糖的测定采用蒽酮比色法(王福钧等,1982;南京农学院,1980);蛋白质的测定采用次氯酸盐比色法(王福钧等,1982);类黄酮的测定采用乙醇浸提法(卢山等,1992;贾之慎等,1995);单宁的测定采用磷钼酸-钨酸钠比色法(王福钧等,1982)。各样品均重复测定3次。
1.4 林下植被调查与参考文献张飞萍等(2004)相同。
1.5 分析方法采用逐步回归分析竹叶化学成分与毛竹叶部主要害虫的关系;采用典型相关性分析竹叶化学成分、林下植被与节肢动物群落的关系(李春喜等,2001;唐启义等,2002)。
2 结果与分析 2.1 不同管理措施下竹叶4类化学成分含量不同管理措施下竹叶4类化学成分含量见表 1。从各化学成分的年总体含量(均值A)看,各管理措施之间大多不具显著差异,仅劈草管理的蛋白质含量显著高于化学除草,单宁含量显著低于撂荒和化学除草;从各化学成分在不同管理措施之间的月均含量(均值B)看,各月基本均以蛋白质含量最高,然后依次为可溶性糖、单宁和类黄酮,但同一化学成分在不同月份之间具有显著差异,总体表现为9月份的可溶性糖和6、7、8月份的蛋白质以及5月份的单宁含量均显著较低,3月份的类黄酮含量则显著较高;从不同管理措施各月各化学成分的含量看,除了9月份各管理措施之间的可溶性糖含量不具显著差异外,其余均存在不同程度的显著差异,但这种差异极为复杂,一些管理措施下同一化学成分的含量时而极高、时而极低,表现为时序变化的不一致,这是经营管理措施、竹叶理化特性和气候、季节等因素综合作用的结果;综上可见,各管理措施对竹叶4类化学成分的年总体含量没有显著影响,但对其月含量及时序变化规律产生了极为复杂而显著的影响,这种影响与毛竹叶部主要害虫的世代变化及季节规律相结合,势必影响着害虫种群乃至群落的时空格局。
|
|
根据毛竹冠层节肢动物的优势害虫组成,以1—6号标准地时间×空间序列上72次竹叶4类化学成分测定的结果和相应时空72次竹缺爪螨(Aponychus corpuzae)、竹刺瘿螨(Aculus bambusae)、南京裂爪螨(Schizotetranychus nanjingensis)、竹小爪螨(Oligonychus urama)、蠕须盾蚧(Kuwanaspis vermiformis)和刚竹毒蛾(Pantana phyllostachysae)的种群数量为基础数据,以竹叶可溶性糖含量(X1)、蛋白质含量(X2)、单宁含量(X3)和类黄酮含量(X4)为自变量,竹刺瘿螨种群数量(Y1)、竹缺爪螨种群数量(Y2)、南京裂爪螨种群数量(Y3)、竹小爪螨种群数量(Y4)、刚竹毒蛾种群数量(Y5)、蠕须盾蚧种群数量(Y6)为因变量,分别进行逐步回归分析,结果见式1)到式6)。
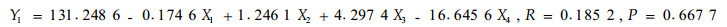
|
(1) |
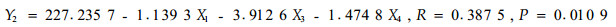
|
(2) |
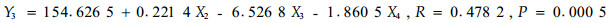
|
(3) |
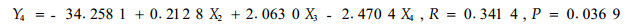
|
(4) |

|
(5) |

|
(6) |
从以上各方程的F检验显著水平和相关系数看,仅(1)式未达显著水平,其余各式均达显著(P<0.05或极显著(P<0.01)水平,各偏相关系数也均达显著或极显著水平,可见式(2)—式(6)为有效方程。式(2)表明,竹缺爪螨与可溶性糖、单宁和类黄酮的关系密切,3类化学成分均抑制竹缺爪螨种群增长,其中单宁的作用最大,然后是类黄酮和可溶性糖;式(3)表明,竹小爪螨与竹叶蛋白质、单宁和类黄酮的关系密切,蛋白质促进该螨种群增长,而单宁和类黄酮起抑制作用,其中单宁的作用最明显,然后是类黄酮和蛋白质;式(4)表明,南京裂爪螨与竹叶蛋白质、单宁和类黄酮的关系密切,蛋白质和单宁均促进该螨种群增长,而类黄酮起抑制作用,其中类黄酮的作用最明显,然后是单宁和蛋白质;式(5)、式(6)表明,蠕须盾蚧和刚竹毒蛾均与蛋白质和类黄酮具密切关系,2类化学成分均抑制两种害虫种群的增长,其中类黄酮的作用大于蛋白质。
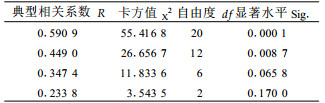
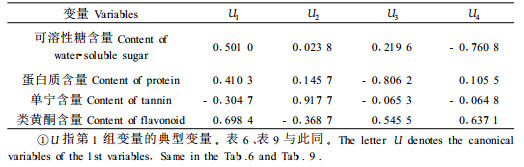
2.3 竹叶化学成分与毛竹冠层节肢动物类群的关系2.2的分析表明,竹叶4类化学成分对冠层优势害虫种群有着显著影响,因此势必通过此影响到整个竹冠层节肢动物类群的物种分布格局。选择1—6号标准地时间×空间序列上72次对竹冠层节肢动物的调查以及相应时空72次生化测定的结果,采用典型相关性分析竹叶化学成分与竹冠层节肢动物的关系。设第1组变量(X)为竹叶4类化学成分含量,命名为生化组,包括可溶性糖(X1)、蛋白质(X2)、单宁(X3)和类黄酮(X4);第2组变量(Y)为竹冠层节肢动物的群落参数,命名为竹冠层节肢动物组,包括丰富度S(Y1)、个体数N(Y2)、Shannon-Wiener多样性指数H′(Y3)、Pielou均匀度指数J(Y4)及Simpson优势集中性指数C(Y5)。分析结果见表 2、表 3和表 4。
|
|
|
|
|
|
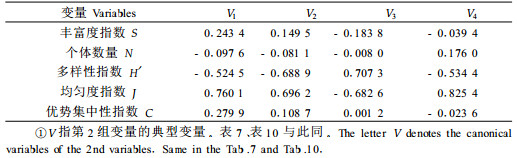
从表 2可知,前2个典型相关系数达到显著水平,可见竹叶化学成分与竹冠层节肢动物类群之间确实存在显著关系。进一步分析第1、2对典型变量,由表 3、4可知,第1对典型变量中,生化组以类黄酮含量载荷最高;节肢动物组以均匀度指数载荷最高;第2对典型变量中,生化组以单宁含量载荷最高;竹冠层节肢动物组也以均匀度指数载荷最高;可见二者之间的关系主要表现为竹叶类黄酮和单宁对竹冠层节肢动物均匀度的促进作用,而这种作用主要通过对冠层优势害虫种群的抑制而形成(见2.2)。
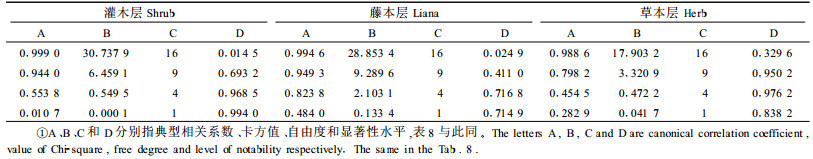
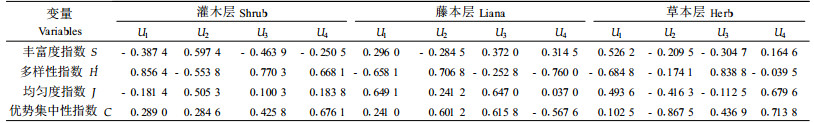
2.4 林下植被与林下节肢动物类群的关系采用对1—9号标准地林下植被的调查数据(张飞萍等,2004)和对1—9号标准地的群落调查数据(张飞萍等,2005a),以标准地为空间重复进行典型相关分析。设林下层节肢动物的群落参数为第1组变量(X),包括丰富度S (X1)、多样性指数H′(X2)、均匀度指数J (X3)和优势集中性指数C (X4),以此为林下层节肢动物组;分别设置林下灌木层、藤本层和草本层植物的群落参数为第2组变量(Y),包括S (Y1)、H′(Y2)、J (Y3)、C (Y4),分别为灌木组、藤本组和草本组。典型相关分析结果见表 5、6、7。
|
|
|
|
|
|

从表 5可知,对于灌木层植物与林下节肢动物的关系,仅第1个典型相关系数达显著水平,由表 6、7可知,第1组变量中以多样性指数H′的载荷最高,第2组变量也以多样性指数H ′的载荷最高,可见2组变量之间的关系主要表现为灌木层植物多样性对林下节肢动物多样性的促进作用;对于藤本层与林下层节肢动物的关系,也由表 5可知,仅第1个典型相关系数达显著水平,由表 6、7可知,第1组变量以均匀度指数J载荷较高,第2组变量以多样性指数H ′载荷较高,可见二者之间的关系主要表现为藤本植物多样性对林下节肢动物均匀度的促进作用。对于草本层与林下节肢动物的关系,所有典型相关系数均未达显著水平。
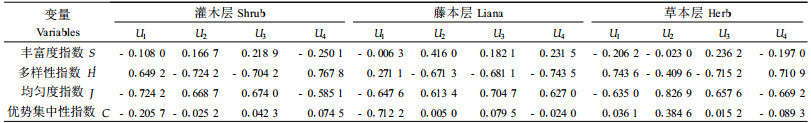
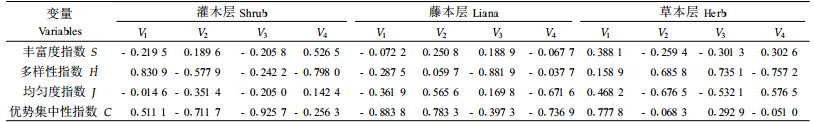
2.5 林下植被与毛竹冠层节肢动物类群的关系林下植被对林下节肢动物的物种分布格局产生了显著影响(见2.4),而林下与冠层的节肢动物在组成与分布方面存在极为密切的联系(张飞萍等,2005b),因此林下植被也同样能够影响竹冠层节肢动物的组成与分布。以同2.4的方法分析林下植被与竹冠层节肢动物的关系,结果见表 8、9、10。由表可知,仅灌木层植物与竹冠层节肢动物的第1个典型相关系数达极显著的水平,且第1组变量以多样性指数H′载荷最高,第2组变量也以多样性指数H ′载荷最高,可见灌木层植物多样性可显著促进竹冠层节肢动物的多样性;藤本层、草本层植物与竹冠层节肢动物的各典型相关系数均未达显著水平。
|
|

|
|
|
|
本文的分析表明:不同管理措施下竹叶化学成分1年中的总体含量大多未表现显著差异,但对其月含量及时序动态产生了极为复杂而显著的影响;这些化学成分与毛竹叶部主要害虫种群的消长具有密切的关系,并因此而影响到节肢动物群落的结构与组成,集中表现为类黄酮和单宁含量显著促进竹冠层节肢动物的均匀度;林下植被也与群落具有密切关系,集中表现为灌木层植物多样性对林下和冠层节肢动物多样性促进作用,以及藤本层植物多样性对林下节肢动物均匀度的促进作用。上述结果说明:经营管理措施可以通过改变林下植被和毛竹生理状况而对群落产生显著影响,不同管理措施下毛竹叶部主要害虫种群数量和群落结构的显著差异(张飞萍等,2005a)也印证了这一点,因此通过科学的经营管理调控竹林生境是害虫防治的有效途径。然而,考虑到野外工作的艰巨性和长期单一措施管理且空间毗邻的对比样地选择较难,本文未能对管理措施设置重复,因而未能进一步分析管理措施、林下植被、竹叶化学成分及节肢动物群落之间的关系,这是本文存在的不足,也是应该继续的工作。
竹叶4类化学成分的年总体含量在不同管理措施之间的差异多不显著,这可能与没有设置重复试验地有关,也可能与野外生境和毛竹生理过程的多样性等有关。但不同管理措施下竹叶4类化学成分的月含量及其时序变化规律具有显著差异,这导致了作为叶部害虫食物的竹叶质和量时序变化的差异,并在气候的时序变化和害虫本身生物学特性的共同作用下,引起害虫种群乃至群落季节变化的差异。本文采用特定时空中竹叶4类化学成分含量与特定时空的害虫种群数量及群落参数进行相关性分析,结果表明4类化学成分均可对某种害虫种群产生显著影响,但对群落的影响则以单宁和类黄酮较为显著,这与群落优势种的组成以及这些优势种与竹叶化学成分的关系等密切相关,同时也说明竹叶中的单宁和类黄酮是影响毛竹叶部害虫的重要物质,这种影响足以显著改变群落的物种分布与多样性水平。
对相邻空间不同管理措施下毛竹林下层植被的研究表明:管理措施对林下植被的组成与分布产生了显著影响(张飞萍等,2004),同一管理措施下在理论上应该具有相近的植被,但在长期的植被恢复过程中,海拔、土壤、林分结构、立竹密度和坡向等因素无疑将产生重要的影响,因此在实践中,不同空间同一管理措施下的植被往往具有较大的差异,这也是笔者在探讨管理措施对林下植被影响一文中(张飞萍等,2004)仅选择空间毗邻的试验地进行比较的主要原因。本文利用9块试验地林下植被的组成与分布情况,分析了林下灌木层、藤本层、草本层植被与节肢动物群落的关系,结果表明灌木层和藤本层与群落具有显著相关性,其中灌木层植被对群落物种多样性的影响最为显著,可见适度保护林下植被多样性,尤其是灌木层的多样性,对于提高群落多样性,增强群落自然控害效能等有着极其重要的意义。
戈峰. 2002. 现代生态学. 北京: 科学出版社, 224-258.
|
贾之慎, 刘志坤, 付一穷. 1995. 竹类中黄酮类化合物总量的研究. 竹子研究汇刊, 14(2): 39-45. |
李正才, 李玉红. 2001. 两种不同类型毛竹林群落特征的初步研究. 竹子研究汇刊, 20(1): 45-49. DOI:10.3969/j.issn.1000-6567.2001.01.009 |
李春喜, 王志和, 王文林. 2001. 生物统计学. 2版. 北京: 科学出版社, 251-254.
|
卢山, 陈晓亚, 夏仲毫. 1992. 方竹属部分种黄酮类成分比较. 竹子研究汇刊, 11(3): 42-47. |
马克平, 刘玉明. 1994. 生物群落多样性的测试方法Ⅸ.多样性的测度方法(下). 多样性的测度方法(下).生物多样性, 2(4): 231-239. |
南京农学院. 1980. 土壤农业化学分析. 北京: 农业出版社, 241-266.
|
钦俊德. 1980. 植食性昆虫食性的生理基础. 昆虫学报, 23(1): 106-121. |
唐启义, 冯明光. 2002. 实用统计分析及其DPS数据处理系统. 北京: 科学出版社, 223-225.
|
王福钧, 严国光. 1982. 农业仪器分析法. 北京: 农业出版社, 266-268.
|
张飞萍, 陈清林, 侯有明, 等. 2004. 毛竹林经营干扰、林下植被及冠层螨类之间的关系. 林业科学, 40(5): 143-150. DOI:10.3321/j.issn:1001-7488.2004.05.024 |
张飞萍, 侯有明, 尤民生. 2005a. 不同管理措施对毛竹林节肢动物群落结构与组成的影响. 昆虫学报, 48(6): 928-934. |
张飞萍, 陈清林, 侯有明, 等. 2005b. 毛竹林冠层与林下层节肢动物类群的关系. 生态学报, 25(10): 2623-2628. |
张飞萍, 陈清林, 吴庆锥, 等. 2005c. 毛竹林节肢动物群落的组成与结构. 生态学报, 25(9): 2272-2283. |
Berenbaum M. 1983. Effects of tannin ingestion on two species of papilionid cate rpillar. Entomol Exp Appl, 34: 245-250. DOI:10.1111/j.1570-7458.1983.tb03329.x |
Chan B G. 1978. Condensed tannin, an antibiotic chemical from Gossypium hirsutum. J Insect Physiol, 24(2): 113-118. DOI:10.1016/0022-1910(78)90106-3 |
Diamond J, 1989. Overview of recent extinctions//Western D, Pearl M C. Conservation for the Twenty-First Century. Oxford: Oxford University Press, 37-41
|
Hedin P A. 1992. Evaluation of flavonoids in Gossypium arboretum cotton as potential source of resistance to tobacco budworm. J Chem Ecol, 18(2): 105-114. |
Klocke J A, Chan B G. 1982. Effects of cotton condensed tannin on feeding and digestion in the cotton pest Heliothis zea. J Insect Physiol, 28(11): 911-915. DOI:10.1016/0022-1910(82)90106-8 |
Schultz J C, Baldwin I T. 1982. Oak leaf quality declines in response to defoliation by Gypsy moth larva. Science, 19: 149-151. |
Waage S K, Hedin P A. 1984. Biologically-active flavoids from Gossypium arboretum. Phytochemistry, 23: 2509-2511. DOI:10.1016/S0031-9422(00)84086-8 |