 2006, Vol. 42
2006, Vol. 42文章信息
- 翟洪波, 李吉跃, 魏晓霞, 徐成立.
- Zhai Hongbo, Li Jiyue, Wei Xiaoxia, Xu Chengli.
- 应用热扩散技术对油松栓皮栎比导率的研究
- Study on Specific Conductivity of Pinus tabulaeformis and Quercus variabilis Using Thermal Dissipation Technology
- 林业科学, 2006, 42(8): 14-18.
- Scientia Silvae Sinicae, 2006, 42(8): 14-18.
-
文章历史
- 收稿日期:2005-04-15
-
作者相关文章


2. 北京林业大学资源与环境学院 北京 100083;
3. 河北省孟滦森林经营管理局 承德 068450
2. College of Resources and Environment, Beijing Forestry University Beijing 100083;
3. Mengluan Forestry Management Bureau, Hebei Province Chengde 068450
在应用热技术(包括热脉冲、热扩散和热平衡)(刘奉觉等,1993;1997;Kramer,1983;Zimmermann,1983)和水力结构理论(李吉跃等,2000;翟洪波,2002;Tyree et al., 1991)之前,如何准确测定出一株树每天到底蒸腾消耗了多少水的问题,曾经长期困扰着林学家和植物生理学家(孙鹏森,2000;翟洪波等,2004a;2004b)。尽管测定森林蒸散的方法很多(Bormann, 1996; Wullschleger et al., 1998; Calder, 1992; Yamanaka, 1998;王彦辉等,2005),但目前只有热技术能够在自然状态下比较准确地连续测定树木的瞬时比导率。本文应用热扩散技术对北京西部山区油松(Pinus tabulaeformis)和栓皮栎(Quercus variabilis)的比导率进行了连续的测定和分析,这些工作有利于从本质上阐明树木的蒸腾耗水问题。
比导率Ks(mL·s-1cm-2或mL·h-1cm-2)是水力结构理论中用来描述植物水分生理特性的参数,定义为单位时间内通过树木单位边材木质部横截面积的流量(Tyree et al., 1991;李吉跃等,2000)。本文用热扩散法测定林分中油松和栓皮栎标准木胸径处的液流速率V(cm·s-1),液流速率是指液流通过树木边材木质部横截面积的流速,与比导率在数值上相等,但意义不同。比导率乘以边材的横截面积即为单位时间通过的液流量F(mL·s -1)。在热扩散法中,由于采用的传感器是线传感器,不同于热脉冲法中的点传感器,其测定的是整个边材木质部导水的平均速率,在计算比导率和液流通量等参数时更加简单方便。如果忽略树体的水分贮存问题,通过树木胸径处断面的水量应该等于树木的蒸腾耗水量,因为根部吸水的99.8%以上消耗在蒸腾作用上(王沙生等,1991),所以根据测定的单木耗水量,可通过边材面积估算到整个林分。可见,标准木胸径处比导率的测定和变化特点,是林分耗水问题的核心。
1 材料与方法 1.1 标准木的选取2001年4—10月,在北京林业大学妙峰山教学实验林场西山燕子岭(海拔410 m)选择油松栓皮栎混交林,林龄31年,林相整齐,郁闭度0.8,林分组成7油3栓,密度1 433株·h m-2。根据每木检尺的结果,选择立地条件和树龄完全相同、位置相邻的油松和栓皮栎标准木各1株,用同一台主机驱动的不同探针(每株标准木南、北两侧各1个探针,取其平均值作为观测值)连续测定其胸径处比导率的变化情况。
1.2 试验方法 1.2.1 比导率的测定选择树干通直、冠形圆满的油松和栓皮栎标准木各1株,用Dynamax公司提供的TDP30热扩散液流探针(Thermal Dissipation Sap Velocity Probe)测定样木的边材液流速率,具体方法见Granier等(1994)所使用的方法。采样间隔与气象因子观测同步,设为10 min 1次,最后用Dynamax公司提供的软件计算边材比导率。
1.2.2 气象因子的观测气象因子观测采用英国Delta-T仪器设备有限公司生产的便携式自动气象站,主要测定的小气候因子包括空气温度、空气相对湿度、土壤温度、降雨量、有效太阳辐射、风速和风向。有关气象站的一些测定参数见表 1。
|
|
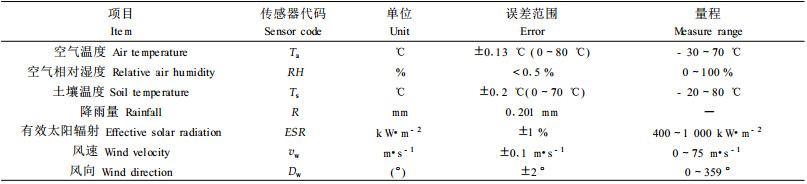
在距2株标准木根茎1 m处埋设土温探头,土温的测定取地表(5 cm)、25 cm、50 cm和70 cm 4层,空气温度和空气湿度的测定分林冠上、林冠中和林下3层测定。
1.3 数据分析所测数据用SAS软件和Microsoft Excel软件来处理。
2 结果与分析 2.1 比导率的日变化特点从整个生长季中任选出一个天气晴朗、光照充足的时间段,来分析其比导率的日变化特点。图 1给出了2001年8月26日8:11到8月28日10:01时段内,混交林中油松和栓皮栎标准木胸径处比导率的连续日变化。从图中不难看出,在自然条件下,油松和栓皮栎的比导率在标准日内均有明显的日变化规律,在早晨6:00左右开始直线增加,在9:00—11:00左右达到最大值,一直到16:00左右基本保持稳定,波动不大,然后开始直线降低,到18:00左右基本降至最低,夜间则几近为0。整个日变化呈明显的凸形变化。由试验的结果可知,比导率在其他月份的标准日内也呈现类似的变化趋势,只是在比较干旱的月份波峰较小,比导率在上午的增加和在下午的减小更趋平缓。阴天时比导率日变化的波峰会明显减小,随着云量的增加,比导率的日变化有时几近直线,但栓皮栎的比导率始终高于油松。

|
图 1 油松栓皮栎标准木胸径处比导率的日变化 Fig. 1 Daily changes of specific conductivity at breast height of P. tabulaeformis and Q.variabilis standard tree 1.栓皮栎Q. variabilis; 2.油松P. tabulaeformis. |
影响树木比导率的因素基本上可以分为2类,即生物学结构的遗传特性和环境因素(包括土壤因子和气象因子)。生物学遗传特性决定了不同树种潜在比导率的大小,这和植物木质部微观结构特征有密切联系,栓皮栎属环孔材树种,其导管直径要远大于油松的管胞直径;环境因素决定了比导率大小的总体水平和瞬时变化。
2.2 环境因子的日变化特点在测定比导率的同时,监测了13项环境因子的连续变化,其采样间隔和比导率的测定完全同步,即每10 min采样1次,这13项环境因子分别是:有效太阳辐射(ESR,kW·m-2)、风速(vw,m·s-1)、风向(Dw,°)、5 cm处土温(T5S,℃)、25 cm处土温(T25S,℃)、50 cm处土温(T50S,℃)、70 cm处土温(T70S,℃)、林内温度(Ta1,℃)、林内湿度(RH1,%)、冠层温度(Ta2,℃)、冠层湿度(RH2,%)、冠上温度(Ta3,℃)、冠上湿度(RH3,%)。
图 2和图 3给出了2001年8月26日8:11到8月28日10:01时段内各环境因子的变化情况。从图中可以看出,在标准日内,有效太阳辐射有比较明显的日变化规律,其变化趋势和比导率的变化趋势基本一致,风速和风向的变化总体上无规律可循,但这并不意味着它们和比导率的变化没有关系。一般来说,风速的增加会加速水汽蒸发过程。当风速较低时,比导率随风速的增加较快,即风速很小的变化都会引起比导率较大的波动,但是大风反而会使比导率降低,因为大风会导致气孔开度降低,甚至关闭。土温的日变化中以5 cm处最为剧烈,其波幅明显高于其他土层;其次是25 cm土层处,其温度变化也比较大;50和70 cm土层处其温度日变化很小,特别是70 cm处,其土温几乎无日变化。土温是通过对土壤中水分的粘滞度、根系生长和吸水能力的作用来间接影响比导率的变化,它的影响不像有效太阳辐射和风速的影响那样直观,但更加持久。不同部位的空气温度在标准日内有明显的日变化规律,并且表现出基本一致的变化趋势,一般是从日出后开始逐渐增加,到正午前后达到最大,然后开始逐渐降低,但在8月份,林冠上的温度>冠层的温度>林内温度;湿度的变化与温度刚好相反,一般是林冠上湿度<冠层湿度<林内湿度,湿度也表现出一定的日变化趋势,一般在日出时较高,以后开始降低,12:00—14:00左右达到最低值,日落前后又开始回升。空气温、湿度通过对气孔开度的作用直接影响树木蒸腾和比导率的变化。

|
图 2 环境因子的日变化 Fig. 2 Daily change of environment factors 1.林内Under the canopy; 2.冠层In the canopy; 3.冠上Above the canopy. |

|
图 3 有效太阳辐射日变化 Fig. 3 Daily change of effective solar radiation |
根据2001年8月26日8:11到8月28日10:01时段内所测定的数据,采用多元线性回归的方法来模拟13个环境因子与油松和栓皮栎标准木胸径处比导率的关系,得如下模型:
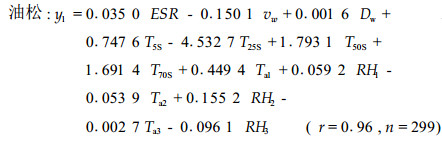
|
(1) |

|
(2) |
式中:y1、y2分别为油松、栓皮栎标准木胸径处的比导率(Ks, mL·h -1cm-2)。可见,尽管各环境因子与比导率的关系未必是线性的,但用线性模型模拟却具有较高的相关性。
用SAS软件进行逐步回归,对每个树种的比导率筛选出3个最重要的预测因子,其统计模型为:
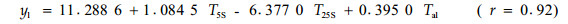
|
(3) |
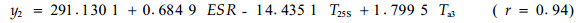
|
(4) |
由软件处理的结果可知,对油松标准木胸径处的比导率影响显著的因子依次为林内温度、25 cm土层土温和5 cm(表土层)土温;而对栓皮栎标准木胸径处的比导率影响显著的因子依次为有效太阳辐射、冠上温度和25 cm处土温。可见,即使在同一林分类似的环境条件下,因为栓皮栎树体高大,占据上层林冠,而油松处于中下林冠层,二者在同一时段内对比导率影响显著的因子差别很大。同一树种在不同时段,由于环境条件的差异,对比导率影响显著的因子也各不相同,比如在9月17—18日和10月13—15日2个时段内的数据,用多元逐步回归筛选3个最重要的预测因子后,可得如下模型:

|
(5) |
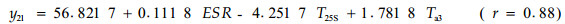
|
(6) |
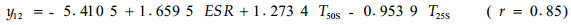
|
(7) |
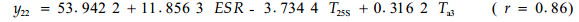
|
(8) |
式中:y11和y21分别为9月17—18日油松和栓皮栎标准木胸径处的比导率(Ks, mL·h-1cm-2);y12和y22分别为10月13—15日油松和栓皮栎标准木胸径处的比导率(Ks, mL·h-1cm-2)。由软件处理的结果可知,在9月17—18日时段内,对油松标准木胸径处的比导率影响显著的因子依次为25 cm土层土温、冠上温度和风速;而对栓皮栎标准木胸径处的比导率影响显著的因子依次为有效太阳辐射、5 cm土层土温和冠上温度。在10月13—15日时段内,对油松标准木胸径处的比导率影响显著的因子依次为有效太阳辐射、25 cm和50 cm土层土温;而对栓皮栎标准木胸径处的比导率影响显著的因子依次为有效太阳辐射、冠上温度和25 cm土层土温。
因此可以肯定,在林分生长期内的不同时段,环境因子对不同树种比导率的作用各不相同,即使是同一树种,在各个时段内对其比导率产生显著影响的环境因子也不是一成不变的,但一般都可以用少数的几个重要环境影响因子来比较准确地模拟出比导率的变化。
3 结论与讨论在野外自然条件下,油松和栓皮栎的比导率在标准日内均呈明显的凸形变化规律,即在早晨6:00左右开始直线增加,在9:00—11:00左右达到最大值,一直到16:00左右基本保持稳定,波动不大,然后开始直线降低,到18:00左右基本降至最低,夜间则几近为0。栓皮栎的比导率始终高于油松,出现这种现象的原因主要是因为栓皮栎是环孔材树种,而油松是针叶(无孔材)树种,树种特性是导致这种试验结果的内在原因。
影响树木比导率的因素基本上可以分为2类,即生物学结构的遗传特性和环境因素。生物学遗传特性决定了不同树种潜在比导率的大小,环境因素决定了比导率大小的总体水平和瞬时变化。这一结论与翟洪波(2002)用油松和栓皮栎苗木进行试验的结果是完全一致的。
在标准日内,有效太阳辐射有比较明显的日变化规律,其变化趋势和比导率的变化趋势基本一致,风速和风向的变化总体上无规律可循,土温的日变化中以5 cm处最为剧烈,其次是25 cm土层,50和70 cm土层处温度日变化很小,特别是70 cm处,其土温几乎无日变化。不同部位的空气温度在标准日内表现出基本一致的变化趋势,一般是从日出后开始逐渐增加,到正午前后达到最大,然后开始逐渐降低,在8月份,林冠上的温度>冠层的温度>林内温度;湿度的变化与温度刚好相反,一般是林冠上湿度<冠层湿度<林内湿度,湿度也表现出一定的日变化趋势,一般在日出时较高,以后开始降低,12:00—14:00左右达到最低值,日落前后又开始回升。
在林分生长期内的不同时段,环境因子对不同树种比导率的作用各不相同,即使是同一树种,在各个时段内对其比导率产生显著影响的环境因子也不是一成不变的,但一般都可以用少数的几个重要环境影响因子来比较准确地模拟出比导率的变化。孙鹏森(2000)用热脉冲技术对油松、栓皮栎标准木进行研究时,也得出过基本一致的结论。
由于干旱半干旱或半湿润地区的土壤水资源不足,林木生长一般都受到不同程度的限制。因此,在研究这类地区土壤水分环境及其蓄水保墒性能的同时,着重研究树木的生理生态耗水特点,有利于最大限度地利用有限的天然降水资源,充分做到适地适树,提高造林成活率、保存率和林木生产力。
李吉跃, 翟洪波. 2000. 木本植物水力结构与抗旱性. 应用生态学报, 11(2): 301-305. DOI:10.3321/j.issn:1001-9332.2000.02.037 |
刘奉觉, 郑世锴, 巨关升. 1997. 树木蒸腾耗水测算技术的比较研究. 林业科学, 33(2): 117-126. DOI:10.3321/j.issn:1001-7488.1997.02.003 |
刘奉觉, Edwards W R N, 郑世锴, 等. 1993. 杨树树干液流时空动态研究. 林业科学研究, 6(4): 368-372. DOI:10.3321/j.issn:1001-1498.1993.04.018 |
孙鹏森.2000.京北水源保护林格局及不同尺度树种耗水特性研究.北京林业大学博士学位论文
|
王沙生, 高荣孚, 吴贯明. 1991. 植物生理学. 北京: 中国林业出版社.
|
王彦辉, 熊伟, 于澎涛. 2005. "多树水分平衡法"的方法与应用. 林业科学, 41(4): 184-188. |
翟洪波, 李吉跃, Huang Wending, 等. 2004a. SPAC中油松栓皮栎混交林水分特征与气体交换. 北京林业大学学报, 26(1): 30-34. |
翟洪波, 李吉跃, 聂力水. 2004b. 油松栓皮栎混交林林地蒸散和水量平衡研究. 北京林业大学学报, 26(2): 48-51. |
翟洪波.2002.中国北方主要造林树种水力结构研究.北京林业大学博士学位论文 http://www.wanfangdata.com.cn/details/detail.do?_type=degree&id=Y480443
|
Bormann H. 1996. Effects of data availability on estimation of evapotranspiration. Physics and Chemistry of the Earth, 21(3): 171-175. |
Calder I R. 1992. Water use of eucalyptus—a review//Calder I R, Hall R L, Adlard P G. Growth and water use of forest plantations. West Sussex, England, 167-179
|
Granier A, Anfodillo T, Sabatti M, et al. 1994. Axial and radial water flow in the trunks of oak trees:a quantitative and qualitative analysis. Tree Physiology, 14(12): 1383-1396. DOI:10.1093/treephys/14.12.1383 |
Kramer P J. 1983. Water Relations of Plants. New York: Academic Press LNC.
|
Tyree M T, Ewers F W. 1991. The hydraulic architecture of trees and other woody plants. New Phytol, 119: 345-360. DOI:10.1111/j.1469-8137.1991.tb00035.x |
Wullschleger S, Meinzer F C, Vertessy R A. 1998. A review of whole-plant water use studies in trees. Tree Physiology, 18: 499-512. DOI:10.1093/treephys/18.8-9.499 |
Yamanaka T. 1998. Evapotranspiration beneath the soil surface:some observational evidence and numerical experiments. Hydrological Process, 12(13/14): 2193-2203. |
Zimmermann M H. 1983. Xylem structure and the ascent of sap. Berlin: Springer-Verlag.
|